

父の先見



父の先見
生命の鉱物起源説
紀伊国屋書店 1988
A.G.Cairns-Smith
Genetic Takeover―and the mineral origins of life 1982
[訳]野田春彦・川口啓明
2週間ほど前、大隅良典さんのオートファジー研究がノーベル生理学医学賞を受賞した。以前から噂のあった研究なので、また朝日賞・京都賞・ガードナー国際賞も授与されていたので当然の受賞だったが、この評価はかなり嬉しい。
オートファジーというのは、細胞がみずからのタンパク質を分解して再利用することである。細胞によっては過剰にタンパク質を合成しすぎたり栄養状態が悪化したりすることがあるのだが、こうしたときに細胞が自分で自分のタンパク質をつくりだして食べはじめることから、オートファジー(自食)と名付けられた。大隅さんは細胞がオートファジーするスイッチをオンにする遺伝子を予想した。これは1993年に見つかった。
自食のしくみがおもしろい。細胞の中に膜(メンブレン)があらわれる。その膜がタンパク質やミトコンドリアなどのオルガネラ(細胞内小器官)をとりこんで、ついで分解酵素を含む別のオルガネラのリソソームと融合すると、とりこまれたタンパク質が分解されてアミノ酸となり、細胞はこれを栄養素とする自食作用をしてのけるのだ。
細胞に自食作用があるらしいことは1950年代からわかっていた。ただ分子レベルでの研究はなかなか進まず、大隅さんが独自に根気よく組み立てた研究によってメンブレン・トラフィックがあきらかになり、オートファジー分野がやっと脚光を浴びたのだった。日本では大阪大学のオートファジー学研究グループもかなり深い研究をしつづけている。阪大の吉森保さんが大隅さんの弟子なのである。
けれどもオートファジー論は、少し前までは「細胞自食仮説」としていささか怪しい議論だと言われていた。そういうギョーカイ反応があった。科学にはつねにこうした紆余曲折がともなうのだが、ここで諦めてはいけなかったのだ。このことは受賞後の会見で大隅さんが少し苦々しい口調で何度も言っている、「流行を追うな、みんながやっているから大事だとはかぎらない、役に立つから科学になるわけではない」と。
若いときに織部や鍋島が好きになると、その後に美濃・楽・黄瀬戸・現代陶芸など気に入ったものはけっこう多くなっていくのに、それはそれ、いつまでたっても織部や鍋島を応援したくなる気分というものがある。ぼくは学生時代にターナーやダリにはまり、またドイツ浪漫派やイタリア未来派にぞっこんになったのだが、いまなおこれらに対する無理解や無定見があると、食ってかかりたいという気分が拭えない。いったん惚れるとずっと擁護したくなってしまうのだ。
科学仮説にもそういうところがある。科学というもの、次々に受容され認定された理論や実験の積み重ねが今日の科学の最前線を構成しているのは当然なのだけれど、とはいえ、さまざまに破れ去った仮説や一部が改変された理論にはいずれもどこか魅力があって、そういう何かの仮説に心が奪われると、その「見方のサイエンス」をずうっと大事にしたくなるものなのだ。
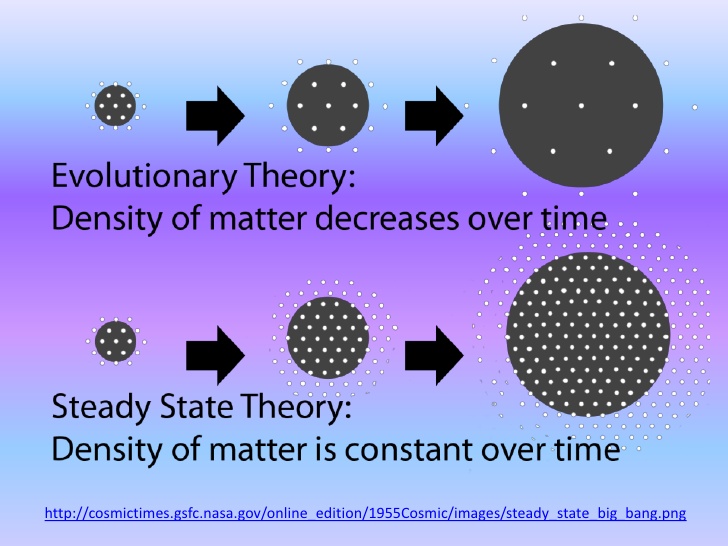
そういうなかには、科学者たちがひそかに捨てられない仮説もけっこう多い。たとえば、天文学におけるフレッド・ホイルの定常宇宙論、生物学における今西錦司の棲み分け理論、数学におけるルネ・トムのカタストロフィ理論などはそういう例だろう。いずれもかなりユニークなものだが、いまは傍流の仮説として放置されている。にもかかわらず、実はこれらが気になっている科学者は少なくない。
それは子供のころ母親の味噌汁や卵焼きに出会ったため、いつまでもその味に郷愁みたいなものを感じることに近いのかもしれない。グラハム・ケアンズ=スミスの「遺伝的乗っ取り」仮説(genetic takeover theory)にも、そうした母の味に似た魅力があった。
生命の起源をめぐる「遺伝的乗っ取り」仮説はいま現在の時点でいうと、その筋書きのすべてが確認されているわけではなく、また生命科学界で了解されているわけでもない。けれども、その大胆で緻密な「読み」はすこぶるスリリングで、いつかこの仮説が多少の変形を受けてもいいからぜひとも稔ってほしいと思ってしまうような、そういうものだった。
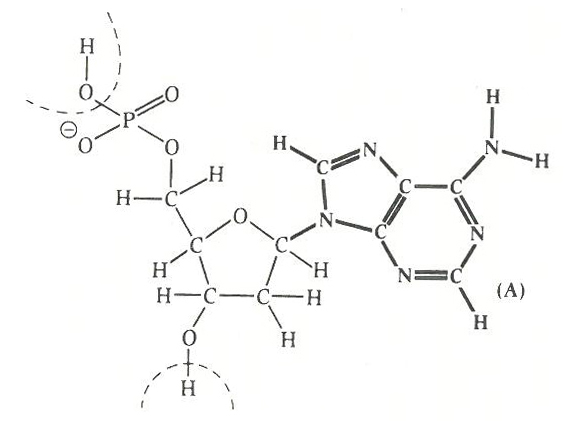
仮説の骨子は、最初の生命体は「ある種の鉱物構造の特徴にもとづいて芽生えたのではないか」ということにある。DNAのような遺伝子が生物体で機能するずっと前、微結晶的なプレ遺伝子のようなものがあって、そのプレ遺伝子が代謝をするようになった初期生命体の細胞の一部を乗っ取って今日の遺伝子になったのではないかというのだ。
鉱物の微結晶性がプレ遺伝子的なはたらきを促したという見方といい、そこに情報を保存したり複製したり編集したりする機能が先行したのではないかという見方といい、ドキッとするような魅力を発揮していた。


生命の起源をめぐる仮説には、古来、さまざまなものがあった。アリストテレスは生物は親から生まれるが、ミツバチやホタルのように草の露から生まれるものがあるとしたし、パラケルススやヘルモントはミミズやカエルやウナギは無生物から発生するとした。
これらは生命の起源を問うというより発生の謎をめぐる発生起源説で、そこには生気(生命の素のようなもの)に形を与える「エンテレキア」(形成力)という謎の力が想定されていた。だからその謎の力にはたいていは無生物との有象無象の関係がひっついていて、いくぶんアニミスティックな憶測がまじっていた。人間にもホモンクルスという“こびと”が原形にいるはずだ、精子の中にはそんな“こびと”の精が宿っているのだろうという憶測も、まことしやかに広まった。
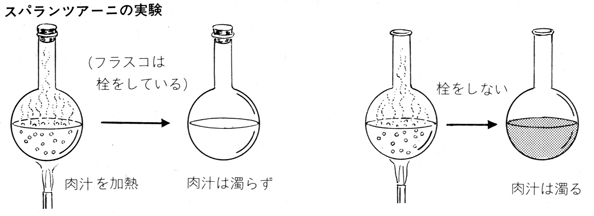
こうした謬見を正面から打ち破ったのはスパランツァーニやパスツールが微生物による発生を説いてからのことで、このとき初めて「無からの発生はない」という見方が確立した。この見方は科学史上ではそうとう画期的なもので、のちのちの「生命科学」や「情報生命科学」の出発点を確立させた。大隅さんではないが、多くの生物発生研究者が細胞や病原菌やタンパク質の動向を探るべく顕微鏡を覗きはじめたのである。
そこへ万能理論ともいうべきダーウィン進化論がかぶさった。生物の分化と進化が自然選択的なプロセスをもつのなら「無生物から生物へ」という脈絡にもこのプロセスがあてはまるだろうというものだ。
こうして物質と生命との、無機と有機との、発生と進化との、それぞれの「あいだ」の脈絡をつなぐ作業が始まった。生命の起源をめぐる議論は「物質から生命へ」「天体要因から地球生命へ」という筋書きを考える科学仮説に向かって、次々に提案がなされることになったのである。
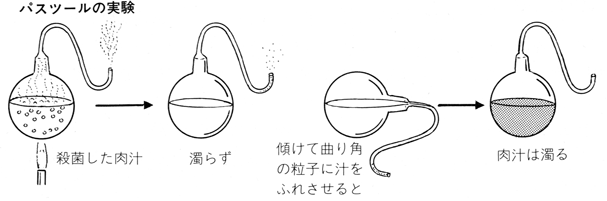
ここではかんたんな紹介にすますけれど、いくつかのアプローチが目立った。まずは化学進化説が走りだした。無機物から有機物が生まれる生化学的なプロセスに立ち入ろうというものだ。スタンリー・ミラーとハロルド・ユーリーの実験が嚆矢となった。水とメタンとアンモニアを含む“原始地球スープ”のミニモデルを容器につくり、これに放電エネルギーを与えたところ有機物が芽生えたというのだ。
ついで解明の対象になったのは代謝機構の解明だった。生命の特質は「代謝」と「遺伝」と「複雑さ」にあるのだが、物質界のどこで代謝が始まったのかに次の研究が向かったのである。早くに名乗りをあげたのが表面代謝説だった。ジョン・バナールやヴェヒター・スホイザーは黄鉄鉱などの表面で有機物の重合がおこって代謝のしくみが生まれたのではないかという仮説を発表した。
一方、生命の起源が海にあるだろうことは見当がついていたので、すぐに海洋生物学が発達した。海のどこかで「酸素をつくった奴」がいるだろうと想定されたのだ。なかで海底奥深い熱噴射孔に生息するシアノバクテリアが注目され、無酸素状態の地球生命の先行部隊の姿のひとつが見えはじめた。こうなると急速に光合成のメカニズムが研究され、植物生理学が生命の神秘に挑むようになった。それとともにATPやミトコンドリアの性質も見えてきた。まとめて「化学合成独立栄養生物仮説」などといわれる。
パンスペルミア説もいくつも提出された。生命の起源は海の底からではなく、天体から生命の種がばらまかれたのだというもので、胚種広布説とか宇宙播種説とかといわれる。フレッド・ホイルやチャンドラ・ウィックラマシンゲやフランシス・クリックがこの仮説を発展させた。
しかし圧倒的な勢いで浮上してきたのは、なんといっても分子生物学と分子遺伝学による成果の数々だったのである。「代謝」と「遺伝」という2つの謎のうちの、いよいよ「遺伝」のほうに研究と仮説が殺到した。
いまでもよく憶えているが、ぼくが「遊」を創刊してしばらくたった70年代半ば、どんな研究所でも大腸菌がもてはやされていた。分子観察対象として大腸菌が脚光を浴びたのだ(いまでも研究者の卵はここから始める)。京大の若き本庶佑さんがキラキラ眩かったころだ。そのうち解析にコンピュータが用いられるようになり、遺伝情報とゲノム情報をデシタル解析するようになった。
分子生物学の成果はめざましかった。予想外の仮説も次々に登場した。たちまち、それまでほとんど顧みられていなかった「情報としての生命」という見方が一貫したシナリオをもちはじめ、これぞ新たな生命観の出現をもたらすだろうと期待された。ぼくはとくにRNAワールド仮説に惹かれた。
けれどもジャック・モノーの話題作『偶然と必然』(みすず書房)やリチャード・ドーキンスの『利己的な遺伝子』(紀伊國屋書店)などを読み継いでいたかぎりでは、残念ながらそこに新たな生命哲学(生命思想)がヴィヴィッドに芽生えたとは思えなかった。とくに遺伝子(つまり遺伝物質)がどのように生きた情報を複製したり編集したりするようになったのか、その根本のところが曖昧だった。そうしたなか、ケアンズ=スミスが「遺伝的乗っ取り」仮説を引っさげて登場したのである。
あらためて仮説の概要を説明すると、最初の生物が出現する以前に、ある種の複製力をもった生命分子のようなものが生じたのだ。それは柔らかい微結晶鉱物の構造のどこかでおこったことで、そこには鉱物の構造そのものの特徴さながらの情報機能が発祥したのであったろう。シュレーディンガーが非周期的結晶性が生命誕生のヒントになりうることを予告していたが、この見方はその延長にあった。
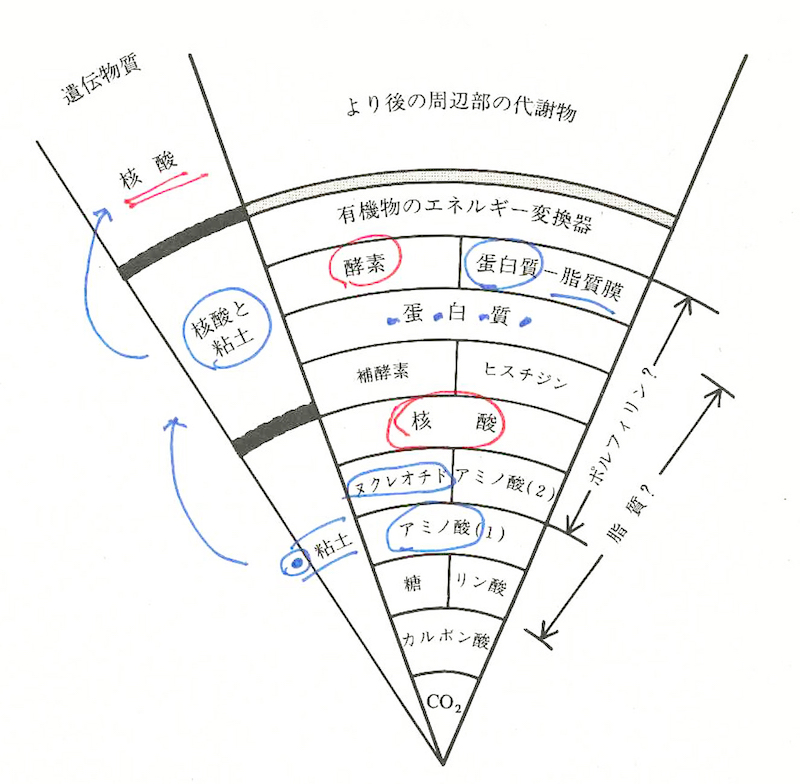
こうしていまだシアノバクテリアが出現していないころ、最初の生命体らしきものが粘土状の鉱物的結晶性を帯びてプリミティブな情報複製能力をもった。これを第一次生命体と呼ぶとすると、この第一次生命体はやがて原初の遺伝現象を示しはじめた。これはいわばプレ遺伝子の動向にあたる。
プレ遺伝子は微結晶性の無機物だが、情報をリプリントしたりコピーしたりする半有機的な能力をつけて、最初の遺伝物質になった。なぜそんなことがおこりえたのかというと、鉱物の結晶の不規則性が対応したのであろうとケアンズ=スミスは考えた。
少しばかり結晶性を帯びたプレ遺伝子(初期遺伝物質)はさまざまな不規則な構造を生じさせるうちに、その欠陥構造ゆえに情報を保存することができただけでなく、自身の自己継承もなしうるようになった。ケアンズ=スミスは、このようなことをおこせたのはカオリンなどの粘土的な鉱物だったろうと想定した。
鉱物の結晶力にはけっこう不安定で不完全なところがある。安定するとはかぎらないし、規則的であるともかぎらない。このことからすると、第一次生命体の原初遺伝子(プレ遺伝子)はもともとはいま知られている核酸やDNAやRNAのようなハイテクなしくみの中にいたわけではなかったと推定される。かなりのローテクだったろう。いやいやゼロテクだったかもしれない。
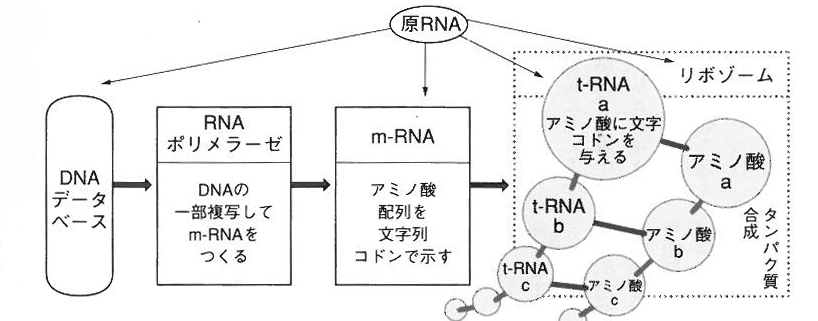
ところが、このローテク遺伝子あるいはゼロテク遺伝子が、次のDNAなどの第二次遺伝子に向かっては、新たな有機的な組み合わせをめざして核酸をつくりだし、情報をもつタンパク質をつくりだしたのである。これは原初遺伝子による「乗っ取り」というべき事件だった。乗っ取りはまんまと成功した。いくぶん結晶的な性質をもった第一次生命体の原初遺伝子は、やがて細胞膜の中に入り込んだ第二次遺伝子としてローテクからちょっとハイテクに機能を上げていったにちがいない。たぶん複製能力を上げたのだ。
こうして、その複製能力やそのことを触媒する能力のある第二次遺伝子がRNAあるいはDNAとなって(第二次遺伝子に進展変化して)、核酸(ヌクレオチド)の中に隙間やらイントロンやらジャンク遺伝子をつくりだし、今日見られるようなDNAによるセントラルドグマに達したのだったろう。
ケアンズ=スミスの専門は物理学および鉱物学である。物質の運動や現象を観察してそこにひそむ変化の具合を読み取り、それを数式に落としこんだり、実験を加えたり、いまだ確定されていない未知の物質を想定したり、そのふるまいを物理的に記述したりすることを得意とする。
しかし、生命の起源のような問題にはなかなかこのような方法があてはまらない。そもそも分野が違う。物理学は無機を扱い、生物学は有機を扱っている。それなら物理学は生命の起源にお手上げかというと、そんなことはない。地球史があきらかにしてきたように、最初の生命は必ずや無機物の組み合わせのどこかから始まっていたはずで、そうであるのなら「物質から生命へ」あるいは「物理から生物へ」という最も神秘なプロセスには、少なくとも半分は物理学者がかかわるべきなのである。
この「物理から生物へ」という最初の一歩は、シュレーディンガーの勇敢な踏み込みとして、『生命とは何か』(岩波新書)という1冊に記念される。そこにはアミノ酸やタンパク質や核酸といった用語にまじって、熱力学や結晶理論の物理用語が入っていた。この踏み込みはとりわけ「生命は負のエントロピーを食べている」「生命は非周期的な結晶作用に似ているかもしれない」という名言にのこっている。
その後もニールス・ボーアからフリーマン・ダイソンまで、物理学は何度も生命の起源の謎に挑んできた。それぞれ示唆的であり、それぞれ理屈が立っていて、それぞれ擽ったいものではあったが、なかでぼくが母の味を知ってしまったのがケアンズ=スミスのジェネティック・テイクオーバー仮説だったのである。
仮説の根底にあるのは、シアノバクテリアなどの生物的生命体が出現するずっと以前に、おそらくは「物質的な生命の分子状態」もしくは「物質情報的な生命分子っぽいもの」があったはずだろうという見方だ。
その生命分子らしきものは多分に結晶的な性質をもっていた。そうでなければ“構造的模倣”をおこす複製力を生じなかった。シュレーディンガーは非周期的な結晶性に生命の本質の匂いを嗅いでいたが、ケアンズ=スミスは鉱物の結晶構造になんらかの欠陥が生じて、そこに無機が有機に向かう発動があったとみなしたのだった。
先にも書いておいたように、鉱物の結晶はしばしば欠陥をもつ。それが地球製の鉱物や岩石というものだ。たとえば結晶にはオーダー(秩序)があるが、欠陥結晶ではそのオーダーが出たり入ったりする。無機物がつくる出入りだが、どこか有機的な出入りに近い。そこへアミノ酸がタンパク質になっていく動向などの「情報の保存や複製や変異」が捻出される背景ができてきたにちがいない。
このような動向はまだ生命活動ではないけれど、十分に情報活動ではあったわけである。だとすればそのうち、このことを継続的に体現してみせるプレ生命分子やプレ遺伝子が情報生命系の芽生えとして生じてもおかしくはなかった。鉱物結晶にはオーダーがあったのだから、このプレ生命分子たちのどこかにはオーダーを組み立てたりそれを継承したりするしくみがなんらかのかたちで引き継がれたのであろう。これはのちの遺伝記号や遺伝暗号の組み合わせを保持したり組み替えたりするのに、まことにうってつけだったのである。
生命を構成する有機分子には厖大な組み合わせがある。わずか10個程度の原子でもヴァージョンがあり、生体構成分子はこれが100個、1000個になるのだから、その組み合わせはすこぶる多能的になる。
もっと驚くべきは生体制御機構としてはたらく酵素のほうで、小ぶりの酵素ですら約5000個の原子を含む。その酵素の作り方のレシピをもったRNAの情報テープではざっと三万個の原子が約束事を守っている。こういうものが組み合わさっているうちに生命体は有機的な連関状態をつくっていくわけである。
どんな手順でこんなふうになるのかといえば、そのおおざっぱな基本は、①分子操作による、②事前調整による、③自己集合による、というものだ。特別な機能をもつ道具立ては①、生ずる効果や結果を限定したいときは②で、分子たちが寄り集まって高次な機能をつくりあげるときは③をつかう。なかで③の自己集合がプレ生命分子の動向としては重要になった。
物質が自己集合するときは、分子が定常的な熱運動の状態にあるときである。この状態では分子は一瞬たりともじっとしていない。なんとかくっつきあおうとし、可能なかぎりの配置を通過しようとする。そのうえで、想定しうるかぎりの安定を求めて特定配置にたどりつく。これが「結晶」になる。結晶化がおこるには、物質状態の温度は高すぎても低すぎてもダメで、かつ分子濃度が十分に高くなっている必要がある。ただし周囲に多くの別な分子がない状態がいい。そういう条件が満たされてくると、分子たちはさかんにくっつきあおうとし、分子間の引き合う力を発揮する。
けれどもすぐさまくっついてはダメなのだ。分子間にはたらく力は可逆的でなければならず、そうであるがゆえに分子たちは自分の配置と位置を次々にテストして、より安定的な構造(構成)を選び出し、結晶化を遂げるのである。
結晶による構造は未飽和・飽和・過飽和によって変わる。エラーもおこるし、不規則にもなる。そして、この結晶構造の微妙な多様性と不安定性がある種の動的な鋳型となって、おそらくはプレ遺伝子のローテクな代替構造をプリンティングさせたのであったろう。ケアンズ=スミスはそういうことをおこさせたのは、きっと無機粘土質の鉱物だったろうと推断してみせた。
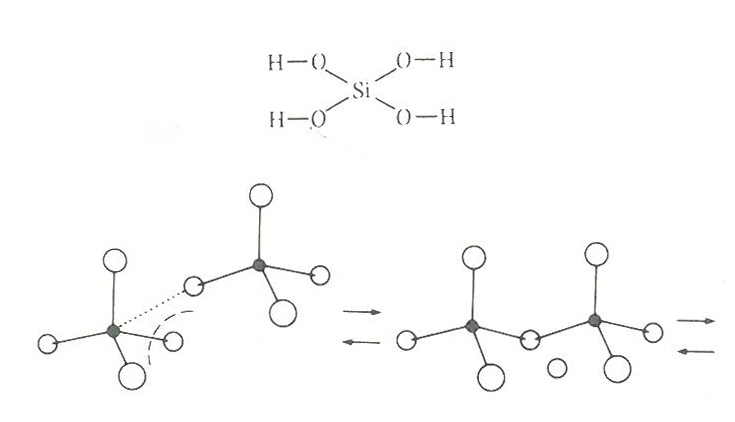
地球は無機粘土質が連続的に結晶化していくプロセスによって成り立っている。粘土は直径0.001ミリほどの粒子でできていて、水には溶けにくいが懸濁しやすい柔岩石である。一つまみの粘土には数種類の無機粘土質が含まれ、その各々に独特の構成単位と結晶構造がある。成分の大半は層状珪酸塩で、その層の出来具合によって2つのタイプに分かれる。
ひとつはカオリンを主成分とする粘土で、この層構造は酸素原子3つぶんの厚さになる。そこでは酸素原子からなる三枚の板が重なっていて、それぞれの隙間で共有結合をおこしている。板の一部はアルミニウム原子でできていて、カオリン層の表面をつくる酸素原子には水素原子が結合する。アルミニウムが介在することはこの結晶に模様を与える。
巨大な粘土層はこのモデルの積み重ねで結晶的理想をめざすのだが、むろんそうはいかないことも多く、そこに不規則性や不連続性もあらわれる。それを鋳型としてプレ遺伝子が育まれたのであったろう。カオリンの変形ヴァージョンにはディッカイトやハロイサイトがあるのだが、これらにはポリティピズム(多型性)が富んでいて、こちらからもさまよう生命分子の可能性が育まれたのかもしれない。
もうひとつの粘土は雲母質のムスコバイトである。大きく整った結晶をもち、対称性が生じることが多い。変形ヴァージョンにイライト、スメクタイトがあり、ここにも初期生命分子が複製力をもっていくためのベッドが用意されていたのかもしれない。
このように粘土質が可塑的であったことが、生命分子の鋳型と逆鋳型の相互作用をもたらし、情報の構造的保存性や継承性をもたらしたのである。それを媒介したのは結晶的遺伝子あるいは半結晶的遺伝子だった。
粘土結晶は規則性と不規則性の両方をもつ。このことは情報をまちがいなく複製すること(すなわち情報のシンタックスを継承しようとすること)、多様な組み合わせによって情報のセマンティックスを生み出そうとすることの、その両方を可能にしたはずである。生命体はこの規則的結晶性と不規則な欠陥構造の両方を生かして先行していったのだ。これ、なかなか魅力的な仮説だった。
ケアンズ=スミスには『生命の起源を解く七つの鍵』(岩波書店)という、やや啓蒙的な本がある。どのように仮説をまとめあげたかという思考のプロセスが述べられていてとても参考になる(かえって紆余曲折をあからさまにしようとして、まわりくどいところもあった)。
その『生命の起源を解く七つの鍵』に、生命の起源を推理するにあたって採るべきいくつかの道標が掲げられている。道具立てについて、生命的統一をめざす結像の条件について、これらを成立させる鋳型の役割についてという、3つの鍵が提供されている。かんたんに紹介しておきたい。
道具立てについては、こうである。
①地球上のあらゆる生命は一揃いの分子だけでなく、共通のシステムをもっていた。その共通のシステムには生化学的な統一性がある。とくにリボソームがタンパク質を合成すること、タンパク質を触媒に用いていること、タンパク質と脂質から生体膜をつくることには、著しい共通システムが動いている。しかしこうした中枢代謝経路の道具立ては生命誕生以前にあったわけではない。生命進化はどの部品を組み立てればいいかという設計図をもっていなかったはずなのである。
②あらゆる生物に共通したシステムには必ずサブシステムが動いている。これらはたいてい連動する。タンパク質は触媒をつくるのに必要だが、タンパク質をつくるには触媒が必要で、核酸はタンパク質をつくるのに必要だが、タンパク質は核酸をつくるのに必要だ。小さい分子たちはそれぞれ相互に依存しあっているのだ。しかしこれらサブシステムは最初から依存しあっていたのではない。ある事態に達してから著しく相互依存するほうに向かったのである。
③共通システムの根本にある特徴は「複雑さ」だ。この「複雑さ」は長期にわたって発揮されたもので、当初の起源的生命体のしくみに備わっていたものとは思えない。
④ということは、共通システムが作動するようになったのは、この生命体にしだいにコンベンション(しきたり)が生まれたからだということになる。すべてのサブシステムもこのコンベンションに従った。そのようなコンベンションが生きたものになっていったのは、この生命体の起源に「型」ないしは「鋳型」があったからである。この「型」にもとづいて先行する生命分子がつくられたにちがいない。
結像の条件については、次のような考え方が用意された。
①究極の祖先が地球で生まれたことはあきらかである。だとすれば、そこには地球が供給した成分がある。
②最初の生命体のための共通システムをつくったのは「自然選択」を促す進化のエンジンであるが、当初のサブシステムには強い相互依存性はなかった。
③強い相互依存性が発揮されたのは、遺伝的乗っ取りがおこってからである。そこにローテクからハイテクへの転換がおこった。
④ローテク生命体は進化することによって、生化学的なハイテク部品を製造できるようになったのであろう。
⑤これらのことをなしとげるために重視されたのは、主には炭素の化学的活用だったにちがいない。
以上のことを怒濤の分化と進化のステージに押し上げる基盤となったのはカオリンなどの粘土質が孕んでいた「鋳型」だった。そこに「無機から有機へ」という大転換事件がおこったのである。
これで「柔らかい鉱物的結晶粘土」に「遺伝的乗っ取り」がおこっていった背景と事情をスケッチしたことになる。本書にはとんでもなく詳細な背景と事情が説明されているので、以上のスケッチではカバーできていないことも多いのだが、それでもここには忘れられない「母の味」がある。
ケアンズ=スミスは、地球上の最初の生物が既存の生命構造からではなく、自然発生的に生じたと考えた。それは第一次生命体といいうるもので、まだ「生きている」とは言えないが、しかしそこから第二次生命体が登場したのだとすれば、このハイテク生命体はそれ以前のローテク生命体のプレ遺伝子が少しずつ異なる遺伝物質を入れ替えていったことによって出現したものなのだ。それは柔らかい鉱物現場における犯人不明の乗っ取り事件だったのである。

⊕ 『遺伝子的乗っ取り―生命の鉱物起源説』⊕
∈ 著者:A.G. ケアンズ・スミス
∈ 訳者:野田春彦・川口啓明
∈ 発行所:紀伊国屋書店
∈ 印刷:精興社
∈ 製本:三水舎
⊂ 1988年 6月24日 初版発行
⊕ 目次情報 ⊕
∈∈ 序
∈∈ 全体の見通し
∈ 1 現在流行している考え方
∈ 2 3つの疑問点
∈ 3 進化に関する疑問
∈ 4 遺伝的乗っ取り
∈ 5 最初の遺伝物質の性質について
∈ 6 最初の生物構成物質
∈ 7 最初の生命
∈ 8 炭素の参入
∈ 9 革命
∈∈ 最終章 問題は将来どのように解かれるか
∈∈∈ 参考文献
∈∈∈ 事項索引
∈∈∈ 人名索引
∈∈∈ 訳者あとがき
⊗ 執筆者略歴 ⊕
A.G. ケアンズ・スミス
1931年スコットランド生まれ、エジンバラ大学卒業、現在、グラスゴー大学科学の講師。1987年より、国際生命の起源学会副会長。本書は生命の起源に関心を寄せる研究者など、関係各方面に衝撃を与え、熱い議論を呼んだ。