

父の先見



父の先見
悠書館 2013
編集:遺伝子図鑑編集委員会・岩井峰人 編集委員長:斎藤成也 序文:太田朋子
装幀:桂川潤
今夜はときどき『銀の匙』を綴った中勘助(31夜)が手元の抽斗(ひきだし)の中に「遺伝子」という未知の代物を見出したというつもりになって、一冊の好ましい図鑑を案内してみたいと思います。でもときどきはオトナになった松岡正剛の言いぶんが入ります。
新町松原を下がった修徳小学校の3階奥の図書室は、司書係のメガネのこわそうなお姉さんをべつにすれば、そこで何をどんなふうに想像したってかまわない「放課後の秘密基地」でした。
勝手にうろうろはできません。本棚の全体を見てまわるなんてことはできないのです(ぼくが図書館や書店をゆっくりまわって見るということができたのは、やっと30代になってからのことでした)。だからあのころは図書室の中を行くたびにずりずりと少しずつカニ歩きをして、見た目に気になる本があると、そこにしゃがんだり坐りこんだりしたのです。
こんな小学校時代の図書室のなかで、夢想のかぎりを遊べたのはなんといっても図鑑のたぐいです。おそらく最初は学研の学習図鑑のようなものを見ていたのだろうと思いますが、そのうち牧野富太郎(171夜)の植物図鑑群にぞっこんになり、あとは百科や大事典や地理図鑑がぞくぞくです。
図鑑に対する興味はその後もあれこれ続き、姉小路東洞院の初音中学時代では学校近くのアメリカ文化センター(ACC)に通ってライフの「自然ライブラリー」シリーズとラルースの絵入り百科を眺めます。
ACCはGHQ(日本を占領したアメリカ軍の総称です)が1952年の日米講和条約発効のあと、全国13都市に開設されたもので、CIF図書館の本が日本の市民が自由に見られるようにしたものです。京都では洋風棟瓦づくりの洒落た建物の中にありました。図鑑がいっぱいでした。ぼくはときどきこの異国情緒に浸っていたのですね。ちなみにぼくは小学校で電気倶楽部に、中学校では科学部に所属していました。
図鑑好きはいまなお体のどこかに届く遠雷のように鳴っています。いつもページを開いているわけではないけれど、突然に見たくなるのが図鑑の魅力というものですから、わがゴートクジの仕事場にもついつい500冊ほどの図鑑系が並ぶようになっています。辞典もの、歴史もの、美術もの、科学もの、百科ものが多いのですが、なかには変な図鑑も混じります。

とはいえ図鑑にも甲乙丙丁があるのです。出来不出来があるのですよ。たとえば、往時の博物学の魂を継承したものもあれば、データのヴィジュアル化に長けたものもあるし、構成と文章はまあまあなのに図解や写真がありきたりで、がっかりするほどひどいものもあります。イラストやポンチ絵が多すぎるもの、やさしい図解解説にしようとしたためにへなちょこフラットになってしまったもの、絵本のような子供用のもの、説明文をただのシェーマに置き換えているだけのもの、そして歴史を飾った博物誌めいた図鑑たち‥‥。困るほどの出来不出来があるのです。
が、総じては、どんな図鑑もチラ見くらいはしておいたほうがいいと思います。ある日ある時、未見の図鑑のページを開いてみるのは知らない星に不時着したときのようなもの、悪かろうはずがないのです。

本書『遺伝子図鑑』は国立遺伝学研究所が4年をかけて仕上げたものです。中身は最新遺伝学に関する「知」とそのヴィジュアライゼーション(見える化)がまんべんなく構成配置されていて、その信頼性はかなり高いものだと思います。
とくに図表化と図解化については、わかりにくい遺伝学の細部のしくみをなんとか一目瞭然にしようと工夫をしています。解説も専門本番用語そのままに使いつつ「ですます調」にしてあるので、遺伝学の入口から出口への道筋がうまく誘導できています。
本文説明もコラム解説も図版説明のキャプションもライターさんが書いたのではなく、すべて研究所の所員たちが分担して書いているので、よかったんですね。これなら十分な筋金入りです。
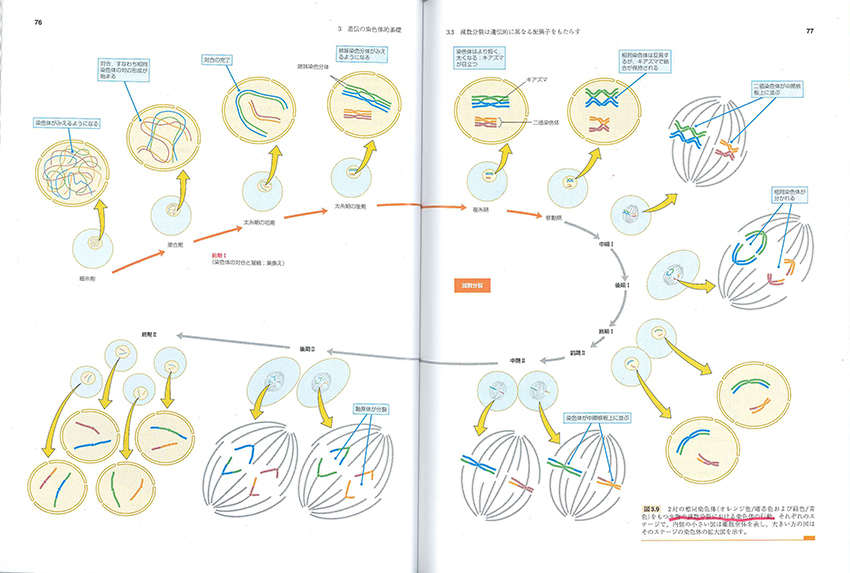
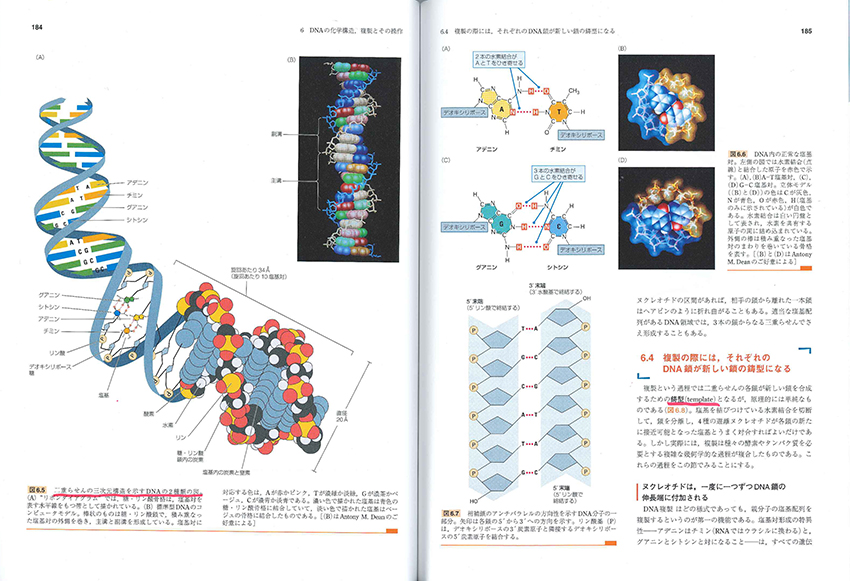
ただしあえて苦言を呈すると、生物的オブジェクトのイラストレーション、その色遣い、模式図の図解表現の仕方、生物学者や遺伝科学者たちの人物画の出来はイマイチです。前夜に紹介したJT生命誌研究館の試みにはとうてい及ばない。ハートルとジョーンズの有名な『エッセンシャル遺伝学』(培風館)の図解性を意識しすぎたのかもしれません。それでも図鑑はすばらしいものです。ウェブなんかより、うんとおもしろい。
この図鑑をつくった国立遺伝学研究所は小ぶりの研究所ですが、日本の生物学系研究所のなかでもなかなかの実績を示してきたところです。
その研究概要はホームページなどで見てください。研究所の専門家たちの成果をもっと読みたいというのなら、参考書としては遺伝学研究所の所員が専門領域を分担執筆した『遺伝子が語る38億年の謎』(悠書館)、いまや研究所を代表するだろう斎藤成也の『遺伝子は35億年の夢を見る』(大和書房)や『ゲノムと進化』(新曜社)などをちょいちょい併読すれば、もっとずっと「遺伝子たちとの格闘ぶり」が立ち上がってくるだろうと思います。



ここから先は遺伝学という学問の進み方についての話を少しずつ入れるので、読み流してもらってかまわないのですが、最近最新の遺伝学(genetics)はそうとう広い領域を占めています。隣接科学も格段にふえている。これは昔日とはほんとうに雲泥の差なのです。
かつては「遺伝のせいだ」というセリフがなんだか重たく、またどこかあやしくて、遺伝に携わる研究といえば暗くて地味なものか、エミール・ゾラ(707夜)のルーゴン・マッカール叢書の計画がそうであったように、「血の宿命」や「家系」や「狂気」に携わっているように誤解されてきたものでした。
この分野のオヤブンの一人の本庶佑(ほんじょ・たすく)さんがどこかで書いていたと思いますが、本庶さんや中西重忠さんが京大の医学部で生命分子の研究に入りかけていたときは、その後の10年ちょっとで分子生物学や遺伝生物学がこんなにもめざましい発展を遂げるだなんてことは、まったく予想できないことだったようです(中西さんは、ぼくのいとこが嫁いだ相手ですが、彼女は急逝してしまいました)。
だから遺伝学はいまなお未曾有の発展を遂げつつある渦中にある学問なのです。当然ながら生物学の一部の学問ですが、最近の遺伝学の進展は次々に生物学全体の深掘りや精査のレベルを押し上げ、推理の翼をどんどん広げる役割を引き受けているように感じられるほど、勢いもあるのです。
その進展ぶりの一部をごくごく雑にいえば、まあ、こんなふうでした。
1950年代後半以降に分子生物学がまたたくまに確立した、DNAやゲノム情報の扱いのオペレーショナルな精度が特段に上がってきた、神経細胞研究や大脳考古学や人類学と遺伝学が近づいてきた、リチャード・ドーキンス(1069夜)の「利己的遺伝子」(selfish gene)の考え方などが広まった、タンパク質研究が広くも深くもなった、ハンチントン病などの遺伝病(優生遺伝疾患)の原因究明が進んだ、ヒトゲノムの塩基配列が短期間で解明された、クローニング技術が成功した、ES細胞やiPS細胞が話題になった、バイオテクノロジーが多くの分野で活用されている、こうしたことが急速な成果をもたらしたので、一挙的な進展がおこったのですね。
最近では「遺伝学的科学思想界」とでもいうべきものが生物学の大きな下敷きになりつつさえあります。

一方、遺伝学という分野そのもののほうも次々に専門的な研究分岐をおこし、ぼくにはほとんどお手上げなのですが、伝達遺伝学、進化遺伝学、集団遺伝学、分子遺伝学、行動遺伝学、発生遺伝学、生態遺伝学、保全遺伝学、ゲノム情報学(ゲノミックス)などなどが独自の成果をあげるようになっています。
それからまた、それらを包んで生化学、進化生物学、分子生物学、数理生物学などが交差したり対比されたり、相互に絡みあったりするようにもなっています。
それだけに、これまで「遺伝」というと学校の生物学でメンデルの法則以上のことを習わなかった人々にとっては、かなり「高度な知」が何機もの戦闘機のように上空を飛行しているということにもなるんですね。幹細胞や万能細胞やiPS細胞のことだって、一般にはかなりわかりにくいことだと思います。
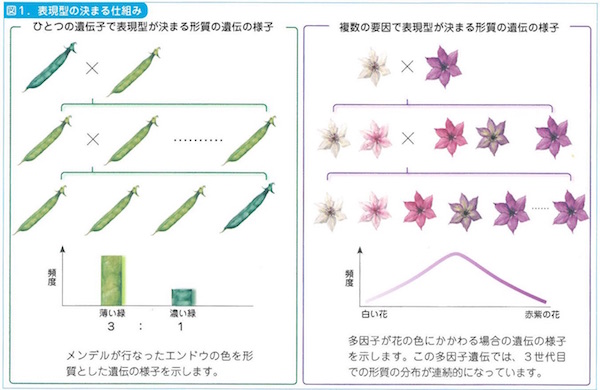
遺伝(heredity)にメンデルの法則が成立していることは、いまでもその通りです。形質(trait)を親から子に受け継いでいくことが遺伝という現象です。
19世紀末、修道院の司祭であって植物フェチでもあったグレゴリー・メンデルがあきらかにしたのは、親の形質がエレメンタルな単位因子(つまりは遺伝子)という単位によって子に伝わっているということでした。
その遺伝にあたっては優性と劣性の力関係があり、優性が発現されると劣性が隠れるという①「優性の法則」がはたらくということ、孫の代になるとその隠れていた劣性の形質が発現してくるという②「分離の法則」があること、優性と劣性とが対立しあう形質はそれぞれ独立して勝手に伝わっていくという③「独立の法則」も成立するということ、この三つの発見がメンデルのもたらしたものでした。ここには、のちのち遺伝子を自立的に扱うという見方がすでに萌芽しています。
ただしメンデルが示したのはエンドウマメの形状や皺の具合にあらわれた7パターンの表現型(phenotype)というもので、遺伝型(genotype)が内部でどういうメカニズムを潜在させていたかはわからなかったのです。メンデルの発見からしばらくたって、ウィーンの病理学者カール・ラントシュタイナーが血液型を発見したときも(例のABO式血液型のことです)、遺伝がもたらす表現型だけが注目されたものです。


その後、モルガンやマラーといった生物学者たちによって伴性遺伝や突然変異のメカニズムがだんだん見えてくるのですが、それでもまだ遺伝の本体はタンパク質が担っているものだと思われていました。
それがアペリー、マクラウド、マクカーティたちの研究で、どうやら遺伝の本体はタンパク質ではなくてDNAが担う化学的遺伝物質にあることが見えてくると、ここから1950年代のことになりますが、オーストリアのエルヴィン・シャルガフらの核酸研究が深まって一気にDNAの存在に脚光が当たり、それが踏み台になってワトソンとクリックによるDNA二重螺旋構造の解明に至ったわけです。
同時期、バーバラ・マクリントックの画期的なトウモロコシ研究によって転移遺伝子(トランスポゾン)も発見されています。遺伝研究はメンデル、シャルガフ、マクリントックの3人で“革命”をおこしたのですよ。
けれども、マクリントックの発見は植物研究(トウモロコシ)の成果だったので、ショウジョウバエやマウスなどの動物中心の遺伝学研究の伝統からすると、彼女に対するノーベル賞(1983)があんなに遅れたのはちょっと理不尽なことでした。なぜか植物派たちは、動物派たちから軽視されていたのですね。ステファノ・マンクーゾの『植物は〈知性〉をもっている』(1613夜)も書いていたことです。
ところでこれらの初期の遺伝学について、ぼくには長らくなじめなかった時期がありました。理由ははっきりしないのですが、たんに不勉強だったか、余計な偏見があったからでしょう(機能主義っぽいなと感じていたのでしょう)。
だからぼくが関心をもったのは、やっとマックス・デルブリュックの業績に馴染んでからのことでした。いまふりかえると、デルブリュックには「物理学から生物学へ、生命科学から感覚生理学へ」という探索の冒険が横溢していたからではないかと思います。ケルン大学に分子遺伝学研究所を、CSH研究所にバクテリオファージ研究所を創設したのもデルブリュックでした。

こうして分子生物学の幕が華々しく切って落とされたのですね。なかでもフランスのジャック・モノーがタンパク質合成の遺伝的制御機構を明らかにすると、遺伝生物学界は大いに盛り上がり、90年代のヒトゲノムの塩基配列解明に向かっていったわけです。モノーの『偶然と必然』(みすず書房)は日本でも学術書としては珍しいほどのベストセラーになりました。
その一方、クローン羊ドリーの話題や遺伝子組み換えの話題が世界を駆けめぐり、はたしてこのままバイオテクノロジー全盛になっていいものか、さまざまな議論も噴出したのです。
遺伝学をめぐってはしばしばこういう毀誉褒貶(きよほうへん)がおこるのです。とくにヒトゲノム計画のゴールが見えてきた1999年、たいへん影響力をもった文明学者のジェレミー・レフキン(824夜)が『バイオ・センチュリー』(集英社)という本で遺伝子技術の産業利用に警告を発し、続いてラジ・パテル(1610夜)の千夜千冊のときに紹介したように、ヴァンダナ・シヴァが痛烈な『バイオパイラシー』(緑風出版)を世に問うと、遺伝子工学や遺伝子組み換え技術がいっせいに非難の的になります。
その後もクリスティン・ドウキンズの『遺伝子戦争』(新評論)、ダニエル・チャールズの『バイテクの支配者』(東洋経済新報社)といった警告が次々に発せられ、「21世紀は遺伝子をいじる時代」だという警戒心も高まったものでした。
日本でも同様のことがおこります。説得力のあるジャーナリストの粥川準二が『人体バイオテクノロジー』(宝島社新書)、『クローン人間』(光文社新書)、『バイオ化する社会』(青土社)などを矢継ぎばやに書くと、「いったい遺伝科学にもとづいた技術って大丈夫なの?」という気運が世間に広まりました。野口勲さんの『タネが危ない』(1608夜)は、そうした危惧から綴られたものでした。

これらのことは必ずしも遺伝学研究の是非が問われたわけではないのですが、そうであればこそ、正確な遺伝についての知識がいまこそ世の中に伝わっていくべきだろうと思います。
さきほどもぼくの認識不足のことを含めて触れておきましたが、遺伝学にはどこか過剰反応や偏見がつきまといます。歴史的にはフランシス・ゴルトンの生殖管理と人種改良を強調した「優生学」(eugenics)の過誤が特段に目立ちますが(それがヒトラー・ナチスのユダヤ人排除のアーリア主義にもなったわけです)、そのほか、いろいろのオーバーランやフライングもある。しかしだからこそ遺伝学の意義をちゃんと知るべきでしょう。
ぼくには21世紀の子供たちはできるだけ早くゲノム情報についての知識に接しておくほうが、英語やプログラムをおぼえることなんかよりもうんと大事なことではないかと思っているので、今夜はちょっとだけ中勘助のふりをしてみたのです。
さて、いまさら言うまでもないことですが、生きものはすべて細胞(cell)からできています。そこには「細胞は細胞からしか生まれない」という根本的な原則が貫いています。一言でいえば、細胞こそが「生命の存在学」を担っているのですね。
その細胞が進化のプロセスのなかでいろいろ変化してきました。それが生物の多様性をつくってきた。なぜ多様化をとげたのかといえば、細胞はいろいろ集まってさまざまな機能を発揮するからです。
細菌や原生生物などの単細胞生物(unicellular organism)では、細胞それ自体がまるごと一個の個体(individual)になっています。一方、真核細胞をスタートとして多様性に挑戦してきた多細胞生物(multicellular organism)では、たくさんの細胞がさまざまな組織(issue)を構成してつくりあげています。私たちヒトもここに属します。ヒトは真核生物なのですよ。このしくみはかなり複雑です。ヒトには約200種類の細胞たちが約60兆近くあって、寄って集(たか)って劇的な組織や器官(organ)を構成しているのです。
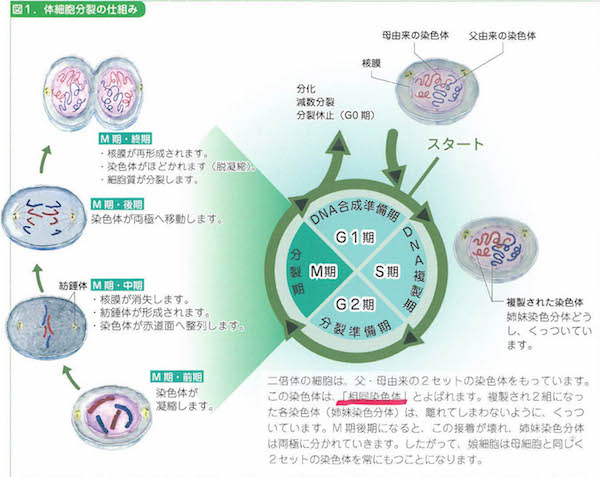
細胞には2種類の細胞があります。受精をおこすための生殖細胞(germm cell)と体をつくるための体細胞(somatic cell)です。体細胞は2セットの、生殖細胞は1セットのゲノムをもちます。ゲノム(genome)というのは、その生物個体がもつすべての遺伝情報のトータルセットのことです。遺伝情報を担うのはそのゲノムの一部を扱う生殖細胞だけで、有糸分裂のような通常分裂(mitosis)か、減数分裂(meiosis)かによって、それぞれの配偶情報を保持します。
こうして生物には細胞周期(cell cycle)が生まれ、それが神の恩寵をうけたかのように維持されてきたのですね。この維持あるいは制御を担当しているのはCDK(サイクリン依存キナーゼ)というタンパク質リン酸化酵素なのですが、数年前から、ぼくはなぜかこのCDKに一目惚れしています。
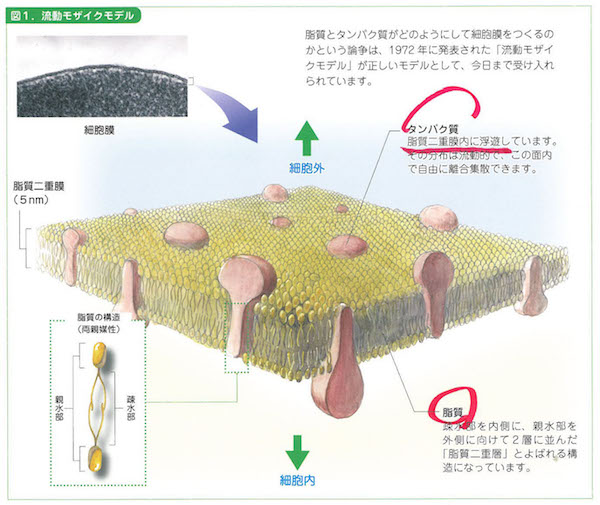
それでは少しだけですが、図鑑の中に入ります。各ページのいろいろの図によって示されているように、細胞を外から観察すると、まずは細胞膜(cell membrsne)が細胞を包んでいることに気付きます。この膜はリン脂質が二重に重なった膜で、生命体を外部と内側に分けているとても重要な膜です。まことによくできたイオンチャネル型の浸透膜です。
ぼくが校長先生をしているイシス編集学校というネット上の学校では、守破離コースの最高難関の「離」コースで、何がなんでも細胞膜(生体膜)のことから編集的生命像の入口を学習してもらっています。11年前からのことでした。そのくらい細胞膜は大事です。
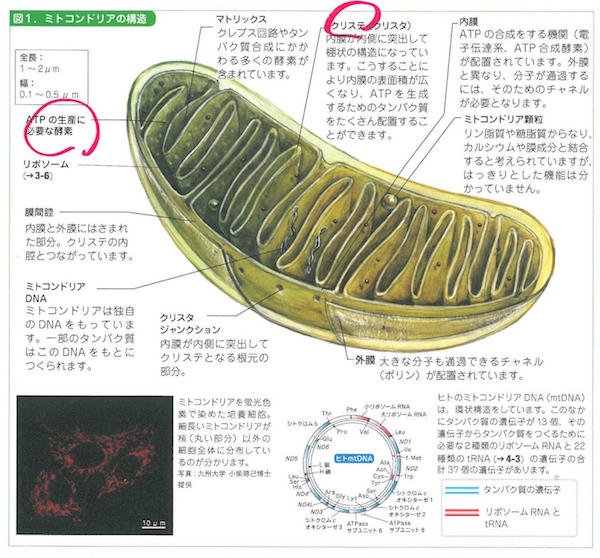
細胞膜の中は細胞質(cytoplasm)になっています。その真ん中付近に核(nucleus)があります。この核の中で遺伝子のやりとりがあるのです。細胞質にはこの核のほかに、リボソームやミトコンドリアやゴルジ体などのオルガネラ(細胞小器官)が入っています。オルガネラという響きには、かのモネラの哲人エルンスト・ヘッケルの香りがします(これはぼくの個人的な印象なので、うっちゃっておいてください)。
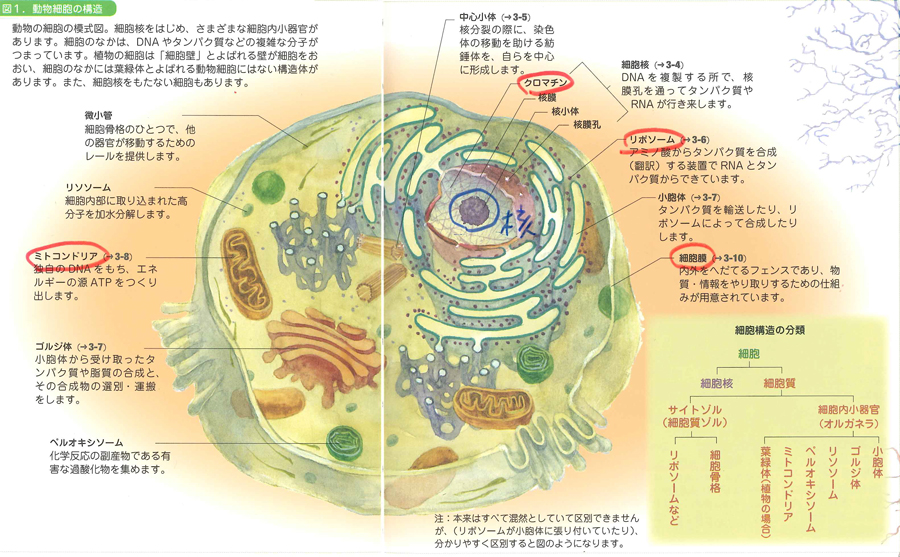

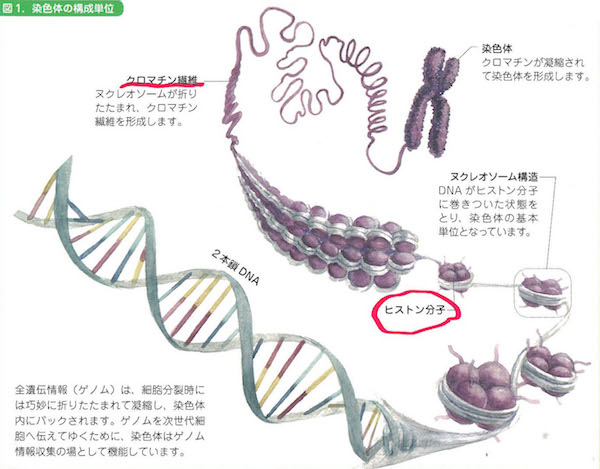
核の中を覗くと、細胞分裂のときに出番を待っていた役者群のように、舞台上に忽然とあらわれる染色体(chromosome)が見えてきます。なぜ役者のように登場したと見えるかというと、染色体はふだんはぼうっとしていて、細胞分裂のときに1万倍くらいぎゅっと凝縮するからです。そうするとシマシマ模様まで見えてくるんですね。
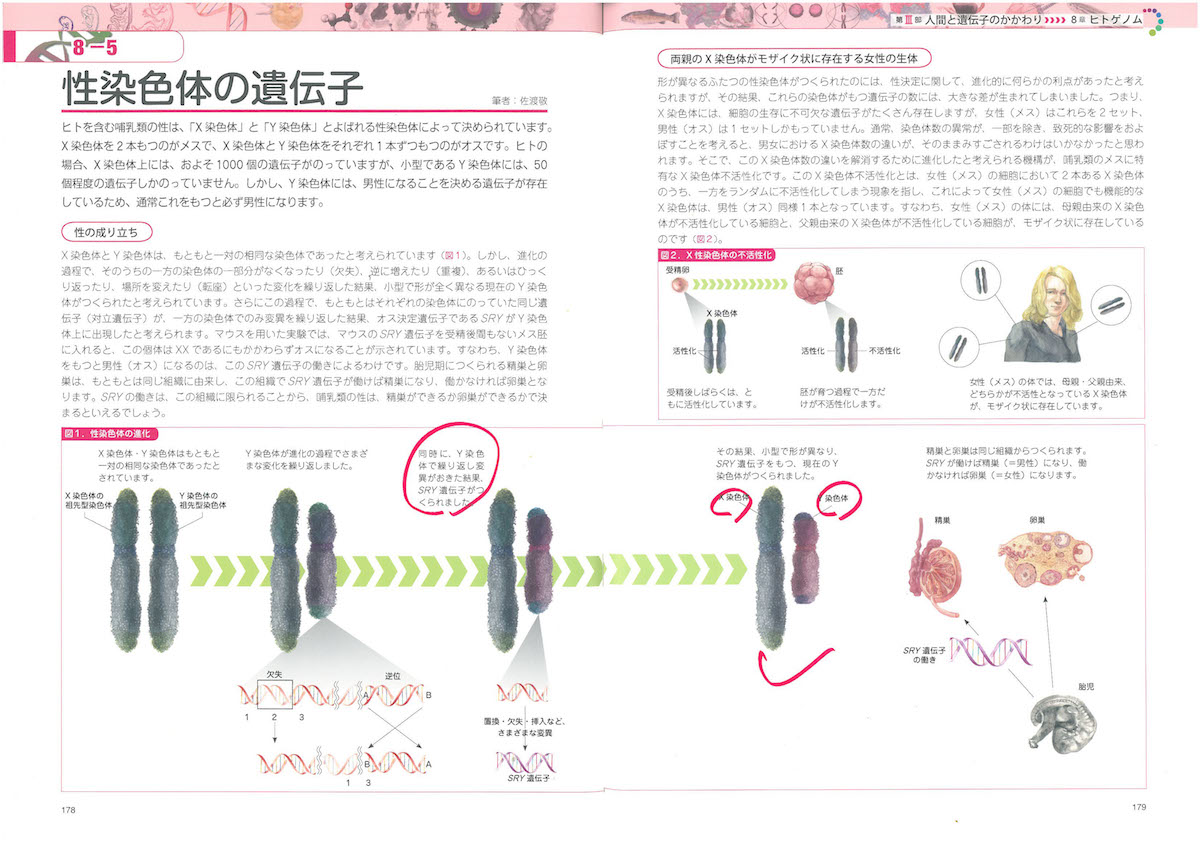
ところで、ヒトの体細胞には46本の染色体があります。常染色体が22本×2で44本、性染色体が2本で、合計46本。
性染色体は女性がXX、男性はXYになっていて、ここにも遺伝子のはたらきが関与しました。もとはXXだったのがオス化遺伝子のSRYがはたらいて、オスとメスに分化したのです。オスは付け足しだったのですね。
染色体はのんべんだらりとしたものではありません。細胞分裂のときに分離するためのくびれた部分のセントロメアと、末端を保護するためのテロメアをもっています。このセントロメアとテロメアの研究もずいぶん進みました。セントロメアは染色体の分配に、テロメアは染色体の端っこのマネージにかかわっているようです。
ついで核の中の染色体が折り畳まれている部分をのばしてみると、細い糸が何かに巻き付いてぐるぐるしているのがわかります。このぐるぐるがDNAそのものです。ついに登場してきましたね。DNAはヒストンというタンパク質に巻き付いてクロマチン構造になっているので、けっこう安定しています(細胞が壊れないかぎり分解しないほどに安定しているのです)。
しかし正確にいうと、染色体の領域すべてが遺伝情報をもっているのではありません。その中のDNAがとびとびに情報をもっていて、遺伝情報をもっている部分とそうでない部分が散らばっているのです。
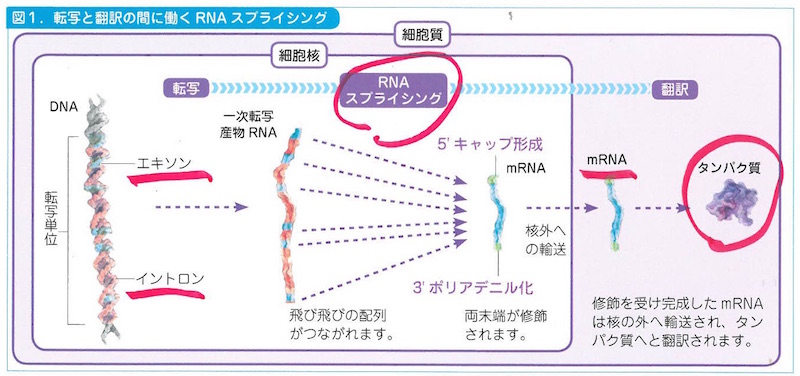
なかでタンパク質についての情報暗号をもっているところがエキソン(exson)で、ここがこのあとタンパク質に翻訳される対象になります。遺伝情報をもっていないところはイントロン(intron)と言って、染色体はこちらのほうが断然に多いのです。最近ではイントロンの役割がいろいろあることがわかってきましたが、長らくイントロンは「空白部」だったのです。だからイントロンはRNAが核から細胞質に出ていくときに巧みにスプライシング(splicing)をおこして除去されます。けれども考えてみれば、これはイントロンにはスプライシングの装置が作動することを識別するシグナルの配列があるということです。イントロンは単なる空白部ではなかったのですね。
このスプライシングのしくみのことを最初に知ったとき、ぼくはやっと分子生物学の凄さを感じたものでした。図鑑5-6「スプライシング」のページを見てください。
しかしなぜイントロンのほうがエキソンより多いのか、その理由はまだわかっていません。宇宙にダークマターが多い理由がわかっていないのと同様、これはかなりの謎ですよ。

以上のことを別の言い方で言い直しますと、染色体は遺伝情報を含んではいるのですが、染色体そのものは遺伝子ではなく、DNAも遺伝子そのものではないのです。DNAのところどころの一部(一画)が遺伝子にあたる、ということになります。
できればこれらはまとめて“遺伝因子”と言ったほうがいいような気がするのですが(そのほうがメンデルこのかたの一貫したジェネティクスが通るように思うのですが)、それはともかくとして、この「染色体>遺伝子>DNA」という離合集散的な相互関係に、前世代から継承されてきた遺伝情報を次世代に伝えるための、いわゆる“設計図”が確実に収納され、その設計にもとづいたタンパク質形成が進行しているのです。
ヒトの例でいうと、ヒトの46本の染色体には約23000の遺伝子が振り分けられているのですが、この遺伝子はヒトゲノムという全体像の中で、それぞれの位置をもっていくのですね。
“設計図”に書かれているのは、どのアミノ酸によってどのようにタンパク質をつくればいいのかということです。そうした設計情報は核の中の染色体の中のDNA遺伝子によって指示されています。その指示にもとづいたタンパク質合成は細胞質のリボソームのほうでおこないます。リボソームは細胞の中のタンパク質合成工場にあたります。
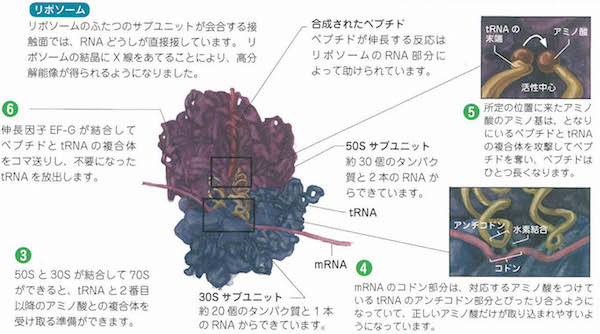
DNAは「デオキシリボ核酸」(DeoxyriboNucleic Acid )のイニシャル略語ですから、本体は核酸(nucleic acid)です。この核酸は4種類のヌクレオチド(nucleotide)が鎖状につながって長い高分子化合物になっていて、遺伝情報の組み立て(設計)を担っています。
核酸はリン酸・糖(オキシリボース=5炭糖)・塩基の3つの要素分子で構成される遺伝物質(genetic material)です。ほかの核酸とちがってかなり特異なハシゴ状の形態と特別な機能をもっています。
構造もはなはだ特異です。糖とリン酸をハシゴの骨組として、そこに塩基のステップが巧みに“上向き・下向き”で向き合い、二重の螺旋階段状態(double helix)をつくっている。これこそがワトソンとクリックが発見した「DNAモデル」でした。
1953年にワトソンらの論文が発表されたときは、DNAは遺伝実態というより担体モデルにすぎないものであったはずですが、驚くべきことにこのモデルは、分子生物学が明らかにするにつれ、まさに生きたモデルとして予測通りの構造としくみをもっていました。凄いですね。
核酸の塩基のハシゴ段の段あるいは手摺りをつくっているのは、A(アデニン)、T(チミン)、G(グアニン)、C(シトシン)という塩基です。DNAはこのATGCという4種類の生命情報記号でできているプログラムの帯束のようなものだと見ればいいと思います。
塩基(base)というのは、もともとは化学用語で「酸」と対になった「塩」(えん)の基体のことですが、遺伝学がいう塩基はA・T・G・Cそのもののことです。
というわけで、ATGCは塩基文字あるいはDNA文字に当たっていて、それぞれの関係には、AとGが水素結合してプリン塩基の、TとCが水素結合してピリミジン塩基の、それぞれの性質を分けもつという相補性がはたらきます。
このように、DNAの情報はたった4種類の文字(生命記号ないしは遺伝暗号)にすぎませんが、これらが一列に順序を変えて次々に単語と文章の関係のように並ぶので、そこにはパターン・ランゲージのようなシークエンス(sequence)があらわれます。
並び順とはいえ、そのうちの3文字ずつが掴めて動かせるのですね。これがトリプレット(3つ組)になったコドン(codon)です。コドンは“単語”の役割をもつので、コドンを組み合わせればかなり複雑な“文意”がいろいろつくれます。コドンは核酸の塩基配列がタンパク質を構成するアミノ酸配列に翻訳されるときの、各アミノ酸に対応しているのです。
けれども、そういう3文字単位のコドンを切った貼ったするというのは、文字や記号や暗号の配列を移したり重ねたりするわけですから、当然、ミスタッチも誤植も脱落もおこります。私たちもキーボードやスマホでミスタッチしてしまいますね。それなら、そんなことがおこると生物は失敗するのかというと、そうでもないのです。失敗も多いのですが、なかにはミスタッチや誤植や脱落は生物の「突然変異」(murtation)として、進化を多様性に富ませていくのです。ここでは「突然変異→アミノ酸の変化→タンパク質の変化→形質の変化」がおきるのです。


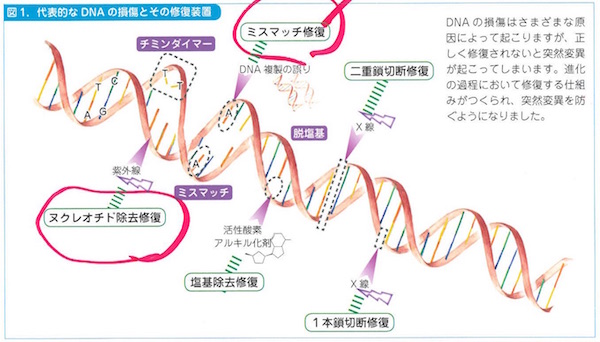
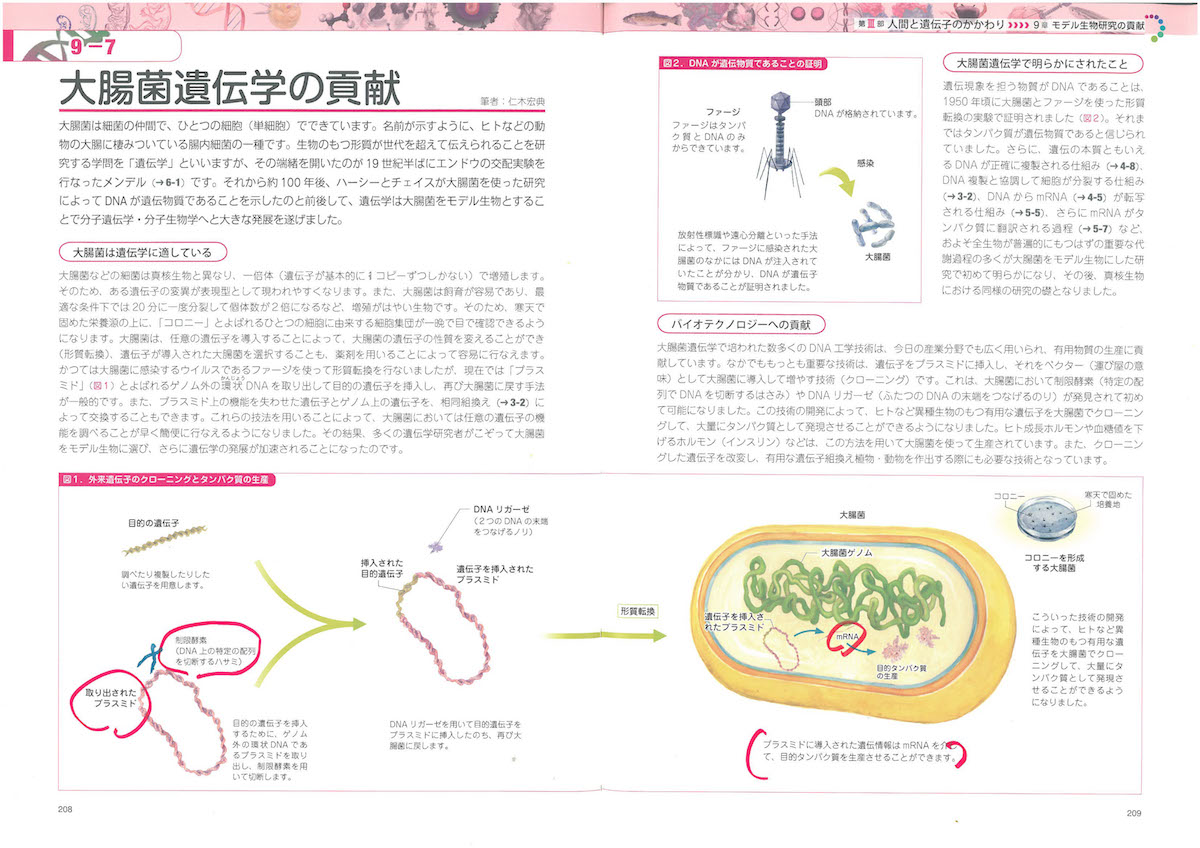
DNAの基本の仕事はまとめていえば「複製、修復、組換え」です。たいへん重要な仕事で、この用語からもわかるように、すこぶる情報編集的なのです。
複製(replication)はコピーができるということです。コピペですね。2個に分裂する細胞が同じ遺伝情報を共有できるようにするためで、DNAが過不足なく倍加します。ここには酵素もはたらいています。
修復(repair)は複製ミスやミスマッチを修復するためのものです。生体分子の多くはディスポーザブルなので(使い捨てっぽいので)、タンパク質やRNAなら遺伝情報があれば合成可能なのですが、DNAはスペアがないので、傷(DNAダメージ)がついたらその場でなんとか復元する必要があるのです。
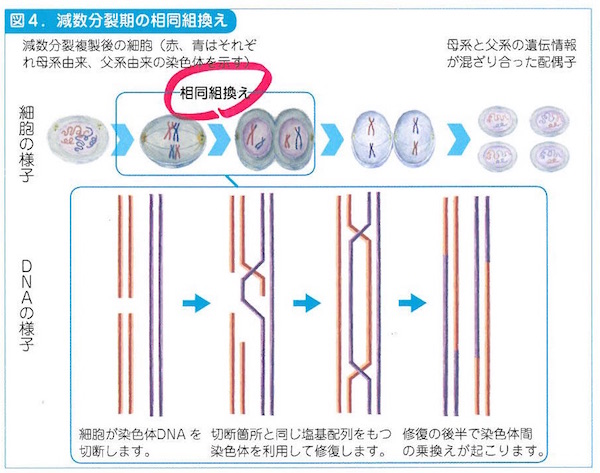
組換え(recombination)は細胞の減数分裂とDNA修復にかかわって補完的におこります。DNA切断で失われた部分を復旧するために無傷の相同配列を借りてくる組換えでは、DNAが部分的に入れ換わります。相同組換えといいます。部位特異的な組換えではリコンビナーゼという酵素がはたらいて、環状DNAがつながって大きくなりすぎることをふせぎます。なかなか巧妙なのです。

生命体が遺伝情報を世代をまたいで継承していく、あるいは変化していくということは、アミノ酸によってタンパク質をつくるしくみを核酸がもっているからです。
遺伝にとってこの最も基本となるプロセスは、遺伝情報が転写され、翻訳されてタンパク質になるというプロセスに集約されます。これによってわずか20種類のアミノ酸によって多様なタンパク質がつくられるということが可能になっているのです。
ただし、ここが重要なところですが、この仕事はDNAだけでは成立しません。もうひとつの核酸であるRNA(リボ核酸)があたかもエディターのように介在するのです。
塩基の配列情報はコドンの組み合わせとして動き出しますが、これはいったんmRNA(メッセンジャーRNA)というRNAに転写(trnascription)されるのです。そのうえでDNAによってタンパク質の言葉に翻訳(translation)されるのですね。
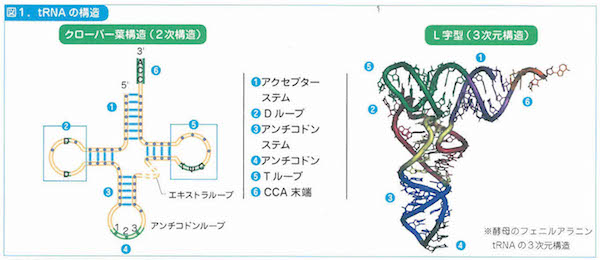
最初の転写では、mRNAがはたらいてイントロンを切り出すRNAスプライシシングがおこります。次の翻訳のプロセスではtRNA(トランスファーRNA)が介在して、タンパク質の折り畳み、輸送、分解が手早く進行します。以上のプロセスがどのように始まって終了するかは、1個の開始コドンと3個の終止コドンが担当します。これらのコドンはストップウォッチをもっているのです。
このような「DNA→(転写)→RNA→(翻訳)→タンパク質」のプロセスこそは遺伝子発現(gene expression)の一番大事なプロセスにあたるので、しかもそれは必ず一方向的に進むので、以前から「セントラルドグマ」と呼ばれてきました。
例外なき王道のようなプロセスだからセントラルドグマと言うのですが、実際にはRNA主導の例外もあります。たとえば、ウィルスのRNA→DNA逆転写、RNA依存のRNA複製、最終産物がタンパク質ではなくRNAであるというケースなどが、そうした例外です。
例外は大事です。基準になる科学がないと何もわからないのですが、でも例外を知ることも、とても大事です。ぼくはどちらかというと、例外がわかると基準の意味がわかるというタチです。
例外のひとつとして、エイズがあります。エイズを発症させるHIV(ヒト免疫不全ウィルス)などのレトロウィルスは、DNAではなくRNAを基体として、逆転写酵素を利用して一本鎖RNAから二本鎖DNAをつくるのです。ぼくはこのことにもびっくりしたものでした。
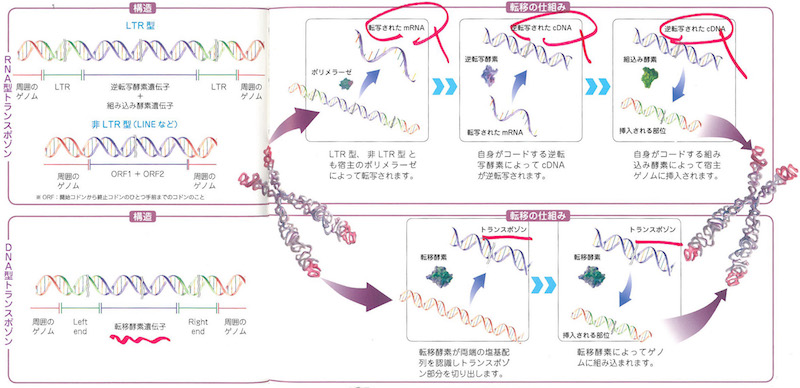
RNAもDNA同様の核酸です。リボース・リン酸・塩基から成っています。ただしDNAのチミン(T)にあたるところがRNAではウラシル(U)に置き換わっているのです。またRNAはDNAにくらべると、水中では化学的に不安定なので、すぐ切れてしまいます。
DNAとRNAにはそういう違いがあるのですが、RNAの特色と機能は必ずしもそのことから生じているのではありません。
これまではDNAこそが遺伝子の主役で、DNAがゲノムの中核を担う因子とみなされていたのですが、DNAをそのようにさせているのは実はRNAではないかという見方があって、この仮説の見地に立つと、 遺伝世界の基盤を用意したのはRNAだったのではないかということになります。
これはDNAが機能する前にRNAが先行的な役割をはたしていたという仮説で、「RNAワールド」仮説といいます。ぼくはずっとこの仮説に惹かれてきました。RNAはきわめて編集的なはたらきを担っている核酸で、生命の起源はこの先行編集によって準備されたのではないかと思っているからです。
とはいえ、RNAワールドがいつどのようにあったのかは確定できません。かなりの太古にRNA型の生命があったのかもしれないし、RNAウィルスのような生命体が何にもまして先行していたのかもしれません。
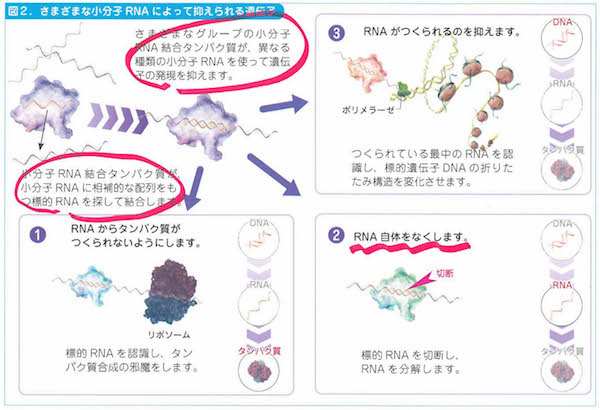
最近になって多くのゲノム領域からRNAが転写されていることがわかってきて、そこにはタンパク質に翻訳されないRNAがはたらいていることが見えてきました。まとめてncRNA(ノンコーディングRNA)といいます。どうやら「備え」にあたっている忍者のようなRNAだろうと思われます。
というような説明をしていくとキリがないのでこのへんにしておきますが、この図鑑にはもっとわかりやすい解説と図示が連打されているので、そちらをこそ一度は覗いてみるのが一番です。きっと中勘助もそうしたでしょう。
本図鑑の構成はⅠ「生物」、Ⅱ「遺伝子」、Ⅲ「人間と遺伝子」になっていて、Ⅰでは生物の多様性、生物のしくみ、細胞の構造とはたらきなどを、Ⅱで遺伝学がもたらした知見の大半と進化遺伝学の概要を、Ⅲでヒトゲノム研究があきらかにしたことの全般と、遺伝病や品種改良などのことが提示されています。全部で11章、108項目。これでわかるように、遺伝子のことだけでなく、生物についての基本の理解ができるようにもなっているのですね。
読者の関心にもよりますが、見逃せない図版構成と解説になっているのは、次のあたりです。これはぼくが勝手に“おすすめ”しているもので、あまり気にしないでください。
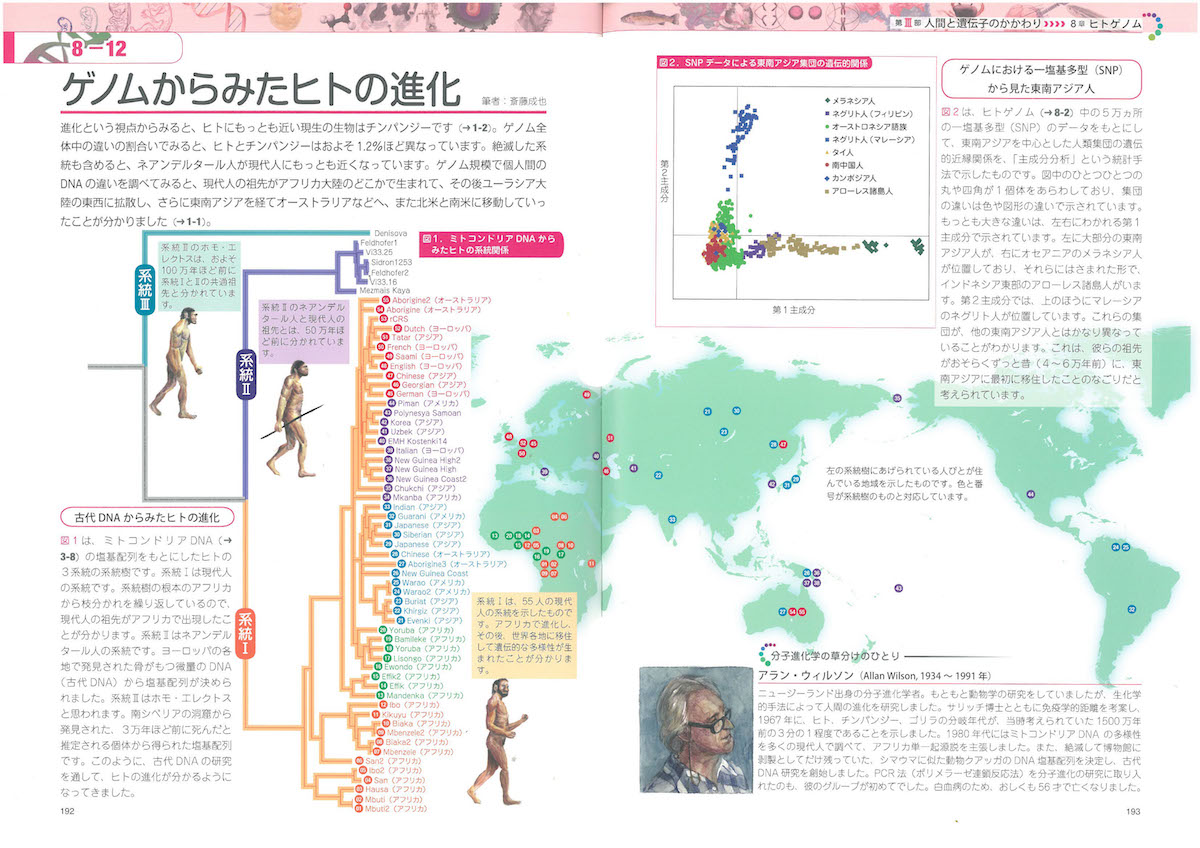
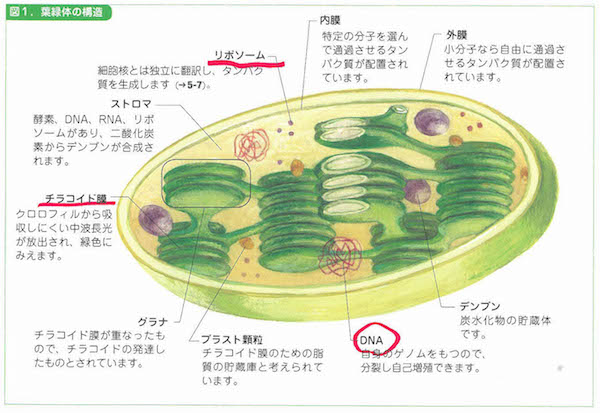
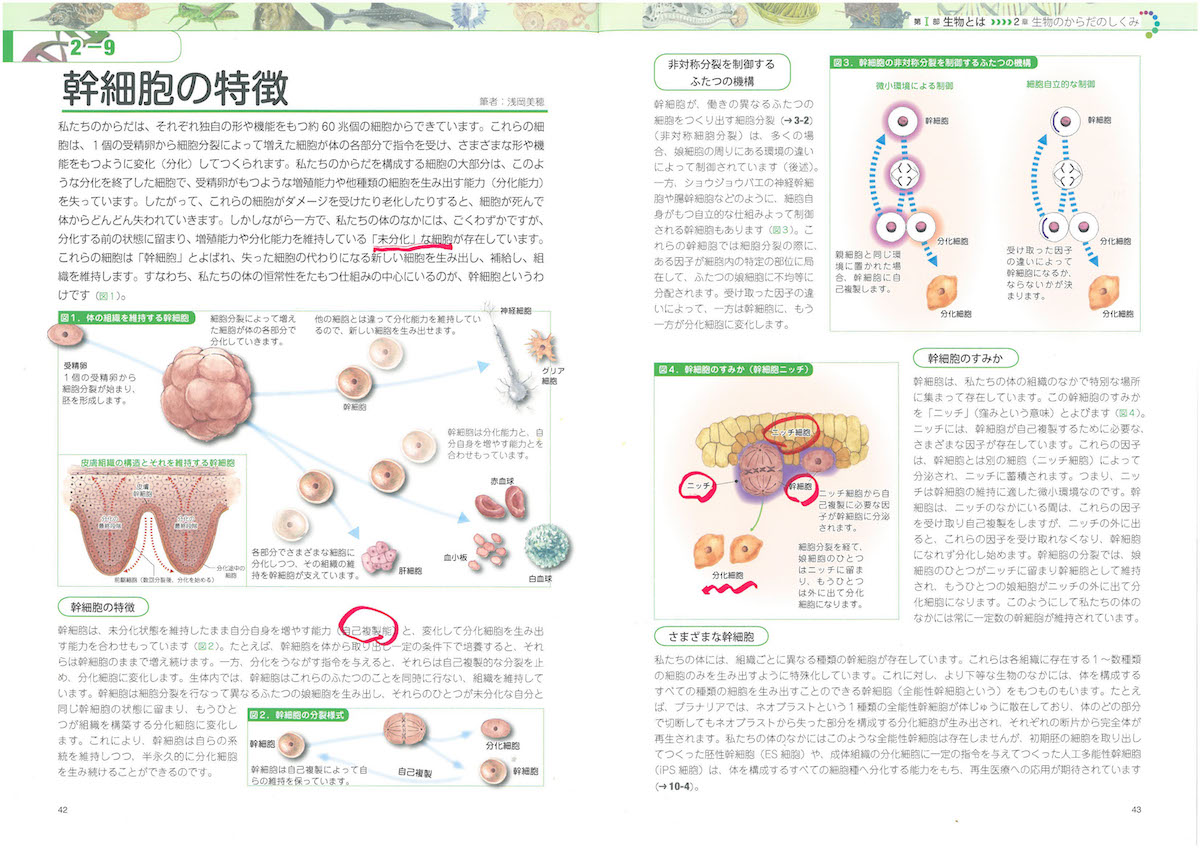
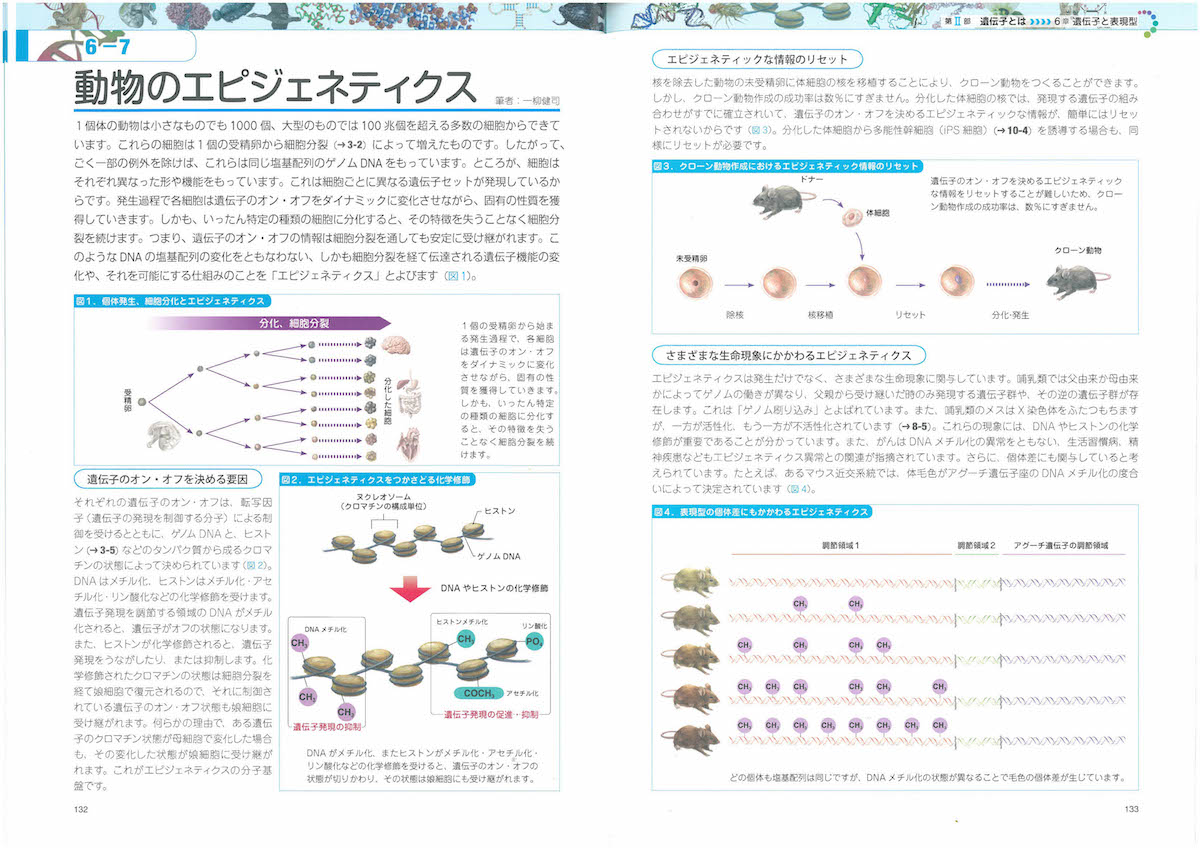
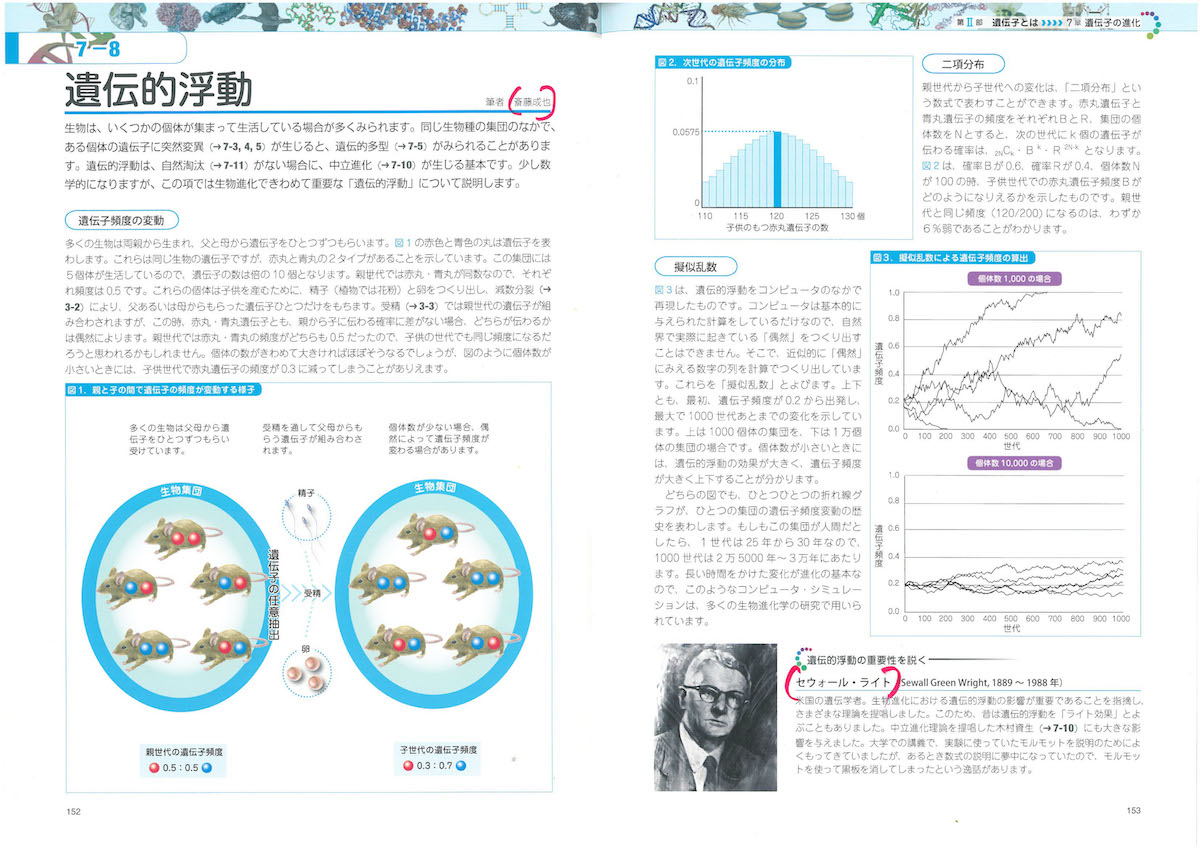
2-9「幹細胞の特徴」、3-2「細胞周期」、3-5「染色体」、3-6「リボソーム」、3-8「ミトコンドリア」、3-9「葉緑体」、3-10「生体膜」、4-3「tRNA」、4-6「小分子RNA、4-9「DNAの修復と組換え」、4-11「動く遺伝子」、5-6「スプライシング」、6-2「表現型と多因子遺伝」、6-7と8「エピジェネティクス」、7-3「突然変異と遺伝的多型」、7-8「遺伝的浮動」、7-13「タンパク質の進化」、8-5「性染色体の遺伝子」、8-10「病気に関連する遺伝子」、8-12「ゲノムからみたヒトの進化」、9-7「大腸菌遺伝学の貢献」、10-5「遺伝子組換え作物」あたり。
ちなみに上記に参考としてあげた国立遺伝学研究所の『遺伝子が語る生命38億年の謎』のほうは、Ⅰ「生物進化の謎」、Ⅱ「人類進化の謎」、Ⅲ「ゲノムの謎」、Ⅳ「細胞と染色体の謎」、Ⅴ「発生と脳の謎」になって、胚性幹細胞(ES細胞)の話で締めくくられています。


ではこれで、図書室の隅っこでうずくまってページをめくってきた「銀の匙」ふうの遺伝子図鑑めくりを閉じることにします。しゃがんでいた腰を伸ばしましょう。
今夜の案内はごくごく入口のところで、図鑑の中身はこんなものではありません。とても詳しいし、とても本格的なのです。生命というもの、つねに本格的なのですよ。生物というもの、いつも驚くべき多様性と例外性に満ちているものなのですよ。でも、その全貌や詳細がわからなくとも、そのどんな部分にも「銀の匙」で一滴すくいたくなるようなキラキラが待っているのです。ここがたまらないところです。諸君もいつか国立遺伝学研究所の一冊を手にとってみてください。

⊕ 『遺伝子図鑑』⊕
∈ 編者:国立遺伝学研究所「遺伝子図鑑」編集委員会
∈ 発行者:長岡正博
∈ 発行所:悠書館
∈ 印刷製本:シナノ印刷
⊂ 2013年 10月25日 初版発行
⊕ 目次情報 ⊕
∈∈ 序文 『遺伝子図鑑』へようこそ
∈ 第1章 生物—その多様性
∈ 第2章 生物のからだのしくみ
∈ 第3章 細胞—生物の構成単位
∈ 第4章 DNAとゲノム
∈ 第5章 遺伝子とタンパク質
∈ 第6章 遺伝子と表現型
∈ 第7章 遺伝子の進化
∈ 第8章 ヒトゲノム
∈ 第9章 モデル生物研究の貢献
∈ 第10章 生活と遺伝子
∈ 第11章 遺伝子の研究方法
∈∈∈ 付録
∈∈∈ 参考文献
∈∈∈ 用語解説
∈∈∈ 索引
⊗ 編者・執筆者略歴 ⊗
国立遺伝学研究所(こくりついでんがくけんきゅうじょ)
1949年、遺伝に関する基礎研究と遺伝学の指導・促進を図ることを目的に、静岡県三島市に設置される。1984年には大学共同利用機関に改組され、現在は総合研究大学院大学の教育も行っている。生命科学分野における中核研究機関として国際水準の先端的研究に取り組む一方、バイオリリース事業、DDBJ(DNA Date Bank of Japan)事業、DNAシーケンシング事業を行っている。
斎藤成也(さいとう・なるや)
国立遺伝学研究所集団遺伝研究部門教授。テキサス大学ヒューストン校生物学医学大学院修了(Ph.D.)。総合研究大学院大学生命科学研究科遺伝学専攻教授、東京大学大学院理学系研究科生物科学専攻教授を兼任。ヒトを中心とする生物の進化をゲノム塩基配列の解析から研究。
荒木弘之(あらき・ひろゆき)
国立遺伝学研究所微生物遺伝研究部門教授。大阪大学大学院理学研究科生理学専攻修了(理学博士)。総合研究大学院大学生命科学研究科遺伝学専攻教授を兼任。出芽酵母を材料として、染色体DNAの複製機構を研究。
角谷徹仁(かくたに・てつじ)
国立遺伝学研究所育種遺伝研究部門教授。京都大学大学院理学研究科博士課程修了(理学博士)。総合研究大学院大学生命科学研究科遺伝学専攻教授、東京大学大学院理学系研究科生物科学専攻教授を兼任。シロイヌナズナという植物を用いて、エピジェネティックな遺伝や反復配列の制御を研究。
小林武彦(こばやし・たけひこ)
国立遺伝学研究所細胞遺伝研究部門教授。九州大学大学院医学系研究科博士課程修了(理学博士)。総合研究大学院大学生命科学研究科遺伝学専攻教授、東京工業大学大学院生命理工学研究科連携教授。細胞の老化と若返りについて、ゲノムの修復の観点から研究。
高野敏行(たかの・としゆき)
京都工芸繊維大学ショウジョウバエ遺伝資源センター教授。九州大学大学院理学研究科博士後期課程修了(理学博士)。国立遺伝学研究所集団遺伝研究部門助教授等を経て、2012年より現職。システムの揺らぎと頑健性、精子形成について研究。