父の先見



父の先見
意識が創られるとき
紀伊国屋書店 2006
本書に出会うまでには少々時間がかかった。こんなふうに言うのは失礼だが、中田力という特異な研究者の仮説の凄みが1冊目ではあまり見えず、2冊目と本書でやっとこさピンときたというところなのだ。
3冊目の本書はライナス・ポーリングの「麻酔に関する先見の明」と「水分子の意味」を切り口として、その脳科学への巧みな理論的応用を説明している。明快だった。切れ味もいい。だが、それ以前の著者の本ではそのあたりの醍醐味が綴られていなかった。いや、ぼくには読みとりにくかった。
本を書くということ、なかなか難しい。アタマの中にあることを全部並べ立てるわけにはいかないし、いったん採用した構成や文脈を変更することもけっこう難しい。それゆえ、そういう本を読むということも、また難しい。しばしば読書における紆余曲折を体験させられる。しかし、読書はそもそもがエディティング・トレーニングなのだから、その紆余曲折にも付き合う必要がある。
できれば、中田さんに会ってから、この紹介を書きたかったのだが、その機会をもてなかった。そのぶん、ぼくがどのように本書にいたる理解の道を紆余曲折したかということを書いておく。そういう書き方もたまにはあっていいだろう。
5、6年前の渋谷ブックファーストでのこと、中田力(なかだ・つとむ)の『いち・たす・いち』(紀伊国屋書店)という本を手にとった。そのころ渋谷ブックファーストの科学書売場は充実していたので、よく立ち寄った。
なんとなく気になる本だということはすぐに感知したのだが、著者の名前とは初お目見え、タイトルもくだけすぎている。なんとかカジュアルな感じにしようとしているようでいて、どうもうまくない。かつヨコ組でもある。ヨコ組は科学書には多いけれど、それにしては専門書の風采ではない。数式がちょくちょく出てくるのでヨコ組にしたのであろう。
残念ながら装幀もよくない。かつて新宿ローヤルマンション1001の工作舎のぼくの近くでスタッフをしていた芦澤泰偉君にしては、冴えない表紙デザインだ(彼はぼくの『空海の夢』の再版装幀をしてくれた)。
サブタイトルは「脳の方程式」。こちらがメインタイトルでもよさそうだ。帯には立花隆の「思ってもみなかった角度から脳科学のパラダイムがひっくり返される」というコピーがついている。何、立花隆?
目次を見たが、すぐには中身が読みとりにくい。ふつうの科学書の構成をとっていないからだろう。エッセイのようでもあるが、しかしその一方で本気科学な気分も伝わってくる。自身の仮説をのべるにあたっての視野が広いのである。口絵にはMRIのカラー写真が並んでいて、なにやら主張している。うーん、どういうふうに読もうかな。
ブックファーストの書棚には同じ著者の『ぷらす・あるふぁ』(紀伊国屋書店)も並んでいて、こちらはサブタイトルが「脳の方程式+α」。あいかわらずタイトルはおもしろくない。やはり芦澤君の表紙で横組だが、帯には「複雑系から脳の渦理論へ」とある。ただし、こちらの目次には「リーマン紀元」「渦理論」「禁断の果実」などとある。えっ、渦理論?
パラパラとページを繰っていくと、全体は手短かな記述になっているようだが、シナプスの可塑性、アセンブリー、アクアポリン、ELDERといったキーワードが飛びこんでくる。ふーむ、これは何やら新仮説らしい。
さっそく両方を買って、1階のカフェで1冊目の『いち・たす・いち』を読み始めた。このカフェはタバコが吸えるので、本を買えば必ず寄った。買ってきた本がつまらなくなると、そのまま荷物を置いて売場に行って別の本を入手して、また戻って読んだものだ。これはぼくがよくやる手で、八重洲ブックセンターの中2階が喫煙可能であった時期は、しょっちゅう中座をしては売場を往復していた(その後、渋谷ブックファーストは閉店、それ以前にカフェが閉店してしまった)。
さて、カフェで雑読したざっとした感じを言うと、日本の科学者にしては文章に切れ味がある。アメリカ帰りだからだろうか。けれどもアメリカどっぷりの利根川進など、およそボケた文章しか書けないままである。利根川にくらべるのは悪いが(中田さんに悪いが)、この著者は何かがうんと冴えている。コーヒーを3杯飲み、キャスターを10本吸った。
何が冴えているかというと、従来のめぼしい科学法則のキモを次々に組み合わせながら、これを非線形の複雑系に出入りするバイナリー・システムにもちこみ、そこから「初期条件に敏感な系」を想定してこれを脳の内部に考えるという方向に、心地よい加速感がある。が、この加速力はどちらかというと科学の構築というより工学的な快感だ。確率、自己形成(自己組織化)、イジング、ユニバーサリティ、相転移、カオス、可塑性といった概念を選び抜いて、一気に「脳の方程式」に向かっている。けれども、やはりのこと、何が「脳の方程式」かはどうも見えてこない。
奥付の上部についている著者紹介によると、中田力は1950年の東京の生まれで、学習院・東大・カリフォルニア大学・スタンフォード大学をへてCaltechで脳神経学の教授をしたあと、いまは新潟大学の統合脳研究センターのセンター長をしている。ファンクショナルMRI(機能的磁気共鳴画像)の世界的研究者であって、すでに各界で新しい科学の提唱者としての呼び声が高いとも紹介されている。なるほど、そういう人か。
そうだとすると、MRIの専門家が「脳の方程式」に挑んだというところが、うん、洒落ている。
中田が「脳の方程式」をあてはめようという領域は大脳皮質の連合野である。以前から「こころ」の中枢だと考えられてきた。
大脳皮質は6層のコラム構造をもっているのだが、それがどのように形成されたのかはあまりわかっていない。それを中田は、ニューロンが6層の皮質構造になるには、細胞が次々に移動してグリア細胞のラジアル線維を伝わってくるのであろうことに注目した。また、そのようにして構造が完成するとグリア細胞とラジアル線維が消失することに注目した。このプロセスは決定論的ではなく、非線形的な自己形成(self organization)がおこっている。
もしそうだとすると、ラジアル線維は熱対流にそってのびて、視床から別ルートでやってきた軸索も熱対流をガイダンスとしてしかるべき部位にまで特異的に到達したということなのだろう。そうであれば、この熱対流は複雑系の法則をあらわしているはずで、そこには“透明な設計図”のようなものが描かれているはずだ。
中田はこの“見えない透明な設計図”を埋めていったようだ。そしてそこにはLGS(Lattice Gas Shell)とでもいうべきものがあると想定し、その基本をELDER(Electron-dense Layer and Dendritic Ramification)と呼ぶことにした。詳細はぼくにはまだ掴めなかったのだが、これはグリア細胞が形成する高電子密度層(electron-dense layer)の形成に関係があるらしい‥‥。
専門用語の解説をべつにすると、ざっとはそういう展望なのだが、なぜ以上のしくみに自己形成(=自己組織化)がおこっていると思ったのかが、『いち・たす・いち』ではわかりにくかった。その説明を最終章だけに詰めようとして、説得力を欠いていた。けれども、何かが気になる。
しばらく中田仮説をほったらかにした。よくあることだ。だいたいぼくは多くの本を平行交差して読むほうなので、著者が一冊にこめたロジックにはよほどの共感があればそのまま埋没するけれど、そうでもないときはしばらくわざと放っておく。そのうちいくつかの関連する視点をぶらさげて蝶々だかハチトリだかのように舞々しながら戻ってきて、その視点をコンディションにあわせて複数動かし、その著者の方面にまた向かっていく。そういうふうにする。
けれども、そういうふうに同一の著者に何度か舞い降りて、それでも裏切られない本というのは、実はそんなに多くない。最近のぼくは、古典やエミール・シオラン(23夜)のアフォリズムのようなものを除くと、吉川幸次郎(1008夜)や野口武彦のものにたびたび舞い戻る。「儒」なるものの日本読みに関心があるからで、それを吉川や野口は裏切ることなく満喫させてくれるからである。
中田の本がそのように感じられたのかどうか、正確には思い出せないのだが、とても重大なことを言おうとしていることだけはわかった。そんなわけで2冊目の『ぷらす・あるふぁ』にとりかかったのは、渋谷ブックファーストの1階カフェを出てから2カ月か3カ月たってのことだった。どんなきっかけで再挑戦する気になったかも忘れたけれど、今度はとても気分よく入っていけた。それとともに著者の狙いや全貌のあらましがやっと見えてきた。
書かれていたのはまさに中田オリジナル仮説ともいうべきもので、それを名付けて渦理論(vortex theory)なのである。うまくかいつまめるかどうか心もとないが、おおざっぱには次のような仮説だった。
脳は大脳と小脳と脳幹でできている。このうちの大脳はニューロンで埋まった大脳皮質(cortex)とニューロン群を結ぶ軸索主体の皮質下白質(white matter)からなっていて、見た目にもグレーがかった皮質とのちがいを見せる。「意識」や「こころ」はこの大脳皮質のなんらかの動向によって生まれると考えられてきた。
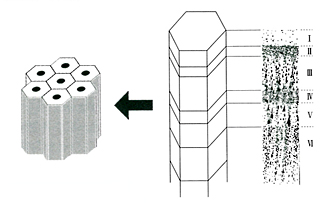
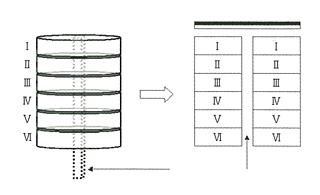
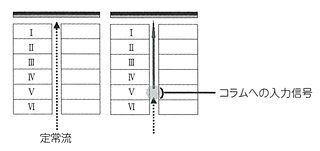
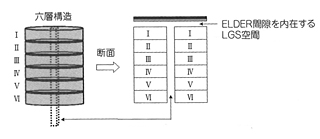
大脳皮質は6層のコラムをユニットとして、これを二次元的に無数に並べたような構造をとっている。鉛筆の束が6本ずつかたまって、それがずらりと並んでいると思えばいい。鉛筆1本が1コラムにあたる。
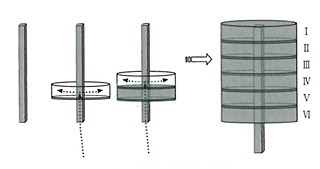
皮質のニューロンが6層コラムとして形成される基本プロセスは、すでに脳科学があきらかにしている。胎生期に骨格構造があらわれ、それが脳の完成時には消滅してしまうのだ。どういうふうにこんなマジックのようなことがおこりうるかというと、皮質を構成するニューロンは自分たちが住みつく皮質で生まれるのではなく、皮質とはかけはなれた神経上皮細胞層で生まれ、そこから皮質コラムに向かって移動する。移動にあたっては、グリア細胞がつくるワイヤーのようなラジアル線維があずかっていて、ニューロンの赤ちゃんをガイドする。そうなっているらしい。

だから、最初に表面まで達した細胞が6番目の層の細胞となり、以降、順次に積み重なっていく。ところがこうして構造が完成するとラジアル線維はもののみごとに消失してしまうのである。あたかも6階建のビルを作るのに、その真ん中に材料を運ぶエレベーターを作っておいて、1階からビルを6階まで作りおわると、エレベーターを取り壊して真ん中をあけ、その後に外部と内部の電線をつないでいるようなものだ。
ここで中田はあることに注目した。ラジアル線維がどのように張りめぐらされているかが、おそらく脳の最終的な形態を決めているだろうということだ。もしもそうなら、その形態形成のしくみこそが大脳皮質の機能を決めている。
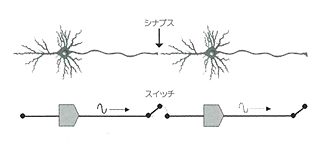
脳はニューロン(神経細胞)によって張りめぐらされている。そのニューロンは樹状突起(dendrite)を入力端子とし、軸索(axon)を出力端子としている。ニューロンの電気的信号は活動電位になっていて、オンするかオフするかの二つの状態しかとらない。入力信号が一定の基準になればオンとなり、次のニューロンに信号が伝わる。ここには悉無律がある。
ただしニューロンとニューロンのあいだにはシナプス(synapse)があって、ここがゲート型のスイッチになっている。このスイッチでシナプスは信号の伝わり方に多少の変化をつける。シナプスには可塑性(シナプス効果)があるわけだ。かつてマカロックとピッツはこのようなニューロンとシナプスの制限的な変化の効果を「重み」(weight)とみなして、有名な神経電子方程式を提案した(NTT出版『情報と文化』参照)。学習は重みの変化でなされるとしたのである。ニューラルネットを議論するときは、この方程式はいまでもよくつかわれている。
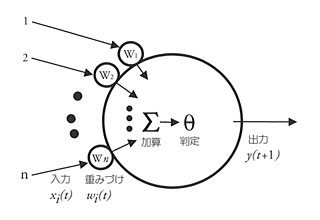
ニューロンに学習性があることは今日の脳科学の大前提になっている。そのため「意識」や「こころ」もすべてニューロンのネットワーク構造だけで説明しようとしてきた。しかし、そのような“ニューロン絶対主義”では説明できないことはいくらもある。マカロックとピッツの方程式だけが「脳の方程式」ではないのだ。
生体はタンパク質、脂質、糖質と金属イオンという、ごくわずかな材料で作られている。その組み合わせから「意識」や「こころ」という情報編集の“極み”を生み出すにはよほどの複雑なしくみが必要だったはずである。とくに人間はここから「ことば」を生み出したのだ。
たとえばDNAはよくできたしくみではあるが、ここには「何を作るか」ということは書かれていない。「どのようにして作るか」というルールだけが書かれている。それなのに、人間はどうして高度な判断や言語や意識をもつようになったのか。この答えはまだ誰も出していないものの、どんなふうにアプローチを組み上げていけばいいのかは、なんとか提案できるかもしれない。
ということで、ここから中田仮説がはじまるのだが、中田はまず機能と形態が合致していくようなマルコフ連鎖による自己形成(自己組織化=self-organization)がおこっていることを前提にした。マルコフ連鎖というのは、現在おかれた状態から次の状態に移行することだけをルールとする情報処理連鎖のことをいう。雪の結晶などもマルコフ連鎖でできている。複雑系の出来事の大半はなんらかのプロセスでマルコフ連鎖によっている。

ラジアル線維が伸びていくプロセスもマルコフ連鎖であるにちがいない。しかもそのことが脳の形態の決定にもつながっている。その形態決定はどのようにおこったのか。マルコフ連鎖だけでは脳は作れまい。中田はここに熱対流(heat convection)が関与していると見た。ラジアル線維は熱対流のルールにしたがっているのではないか。脳の形態は熱対流の全体像と一致しているのではないか。
おそらく熱対流こそが、脳のなかの複雑なしくみを描く「透明な設計図」の何かにあたるのである。そう、想定できる。しかし、これだけでは話はまだまだ半分だ。中田仮説はここからむくむくとアタマを擡げていった。
大脳の形成にあたっては、小脳との比較が有効になる。小脳は出力ニューロンにあたるプルキニエ細胞を中心に構成されている。
プルキニエ細胞は平行線維(parallel cell)から入ってきた多数の入力が一定の閾値に達するとバースト(発火)するようになっているのだが、その伝わり方は一様ではなく、それぞれのシナプス効果にもとづいていく。それによって小脳チップはさまざまな学習ができるようになった。
学習にあたっては、小脳チップが獲得すべき情報とそうでないものがある。それが選択できるからこそ学習なのだが、好ましくない情報(つまりは学習しない情報)については“誤差信号”が出ていると考えられる。プルキニエ細胞はこの誤差信号を、登上線維(climbing fiber)をつかってフィードバックする。登上線維は情報の内容には関与していない。学習すべき内容はシナプスが担当し、登上線維はプルキニエ細胞の学習を促すか中断させるかだけを受け持っている。
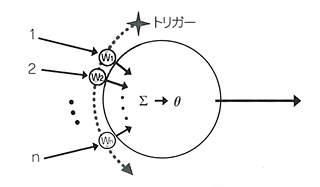
たいそうよくできたしくみになっているのだが、大脳も当然、この小脳のしくみを真似ているはずである。そう、推定したくなる。ところが、大脳には小脳の登上線維にあたるものがまったく見当たらない。何か別のしくみを補完しているようなのだ。
考えてみれば、小脳の構造ですべての情報処理が完了できるなら、脳は小脳から大脳に向かって進化する必要がなかったはずである。もしも容量が足りなかったというのなら、小脳自体が大きくなればよかったのだ。が、そうはなってはいない。大脳は登上線維に代わるしくみをもったようなのだ。そこにこそ大脳に「意識」や「こころ」が生成されたヒントがひそんでいるようだ。
あらためて小脳と大脳をくらべてみる必要がある。小脳のプルキニエ細胞に対応する大脳皮質の出力ニューロンは錐体細胞(pyramidal cell)である。小脳の平行線維にあたるのは皮質内線維で、ここで適度な前処理をうけた結果が錐体細胞に伝えられるようになっている。
ところが大脳には、この錐体細胞に巻きつくはずの登上線維が見当たらない。これがなくては錐体細胞に「さあ、学習をしますか、しませんか」の判断が与えられていないということになる。が、それはおかしい。学習のスイッチはちゃんと入ってくるはずだ。ということは、登上線維の代わりをする何かのしくみがあるということなのである。
そこで中田が組み立てるのは、一元的構造である小脳チップをなんらかの方法で二次元化して、これを大脳が活用したのではないかという予想だった。そうであれば、登上線維のように学習を促す装置がどこかにあって、一度に複数の錐体細胞を刺激できる。そうなれば大脳のなかの三次元構造がつくりあげられる。
どういうものがありうるのだろうか。中田は「分配器」のようなものがあるだろうと睨んだ。
では、それはどこにあるのか。仮に分配器をニューロンで作ってしまえば、錐体細胞に信号を伝えるものがニューロンからの軸索になってしまい、神経線維がダイレクトに伸びることになるが、そんなものは見当たらない。それなら、何が分配器になっているのか。ここでこそ“ニューロン絶対主義”を捨てるべきだろう。
大脳皮質のなかでニューロン以外の細胞を代表しているのはグリア細胞である。グリア(glia)は膠のことで、何かをくっつけているはずである。
そこで中田は、ここには発泡スチロールのようなマトリックス構造めいたものがあるのではないかと考えた。泡構造のようなものがあって、そこに発泡スチロールの空気にあたるような何かが保証されている。そう、読んだのだ。
実は、このマトリックス構造の実在を証明する候補者はすでに見つかっていた。グリアに無数に発見されている「アセンブリー」(assembry)とよばれてきたものだ。近年になってそのアセンブリーの正体は、水の出入りを調節するタンパク質の「アクアポリン4」(aquaporin-4)であることもわかってきた。
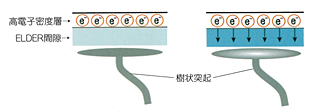
それでどうなっていると考えればいいかというと、グリアはマトリックス構造をつくっていて、それによってそれらの間隙にある水をアセンブリーで吸い上げている。水を吸い上げられた間隙には「乾いた空間」が残る。この空間には電気を通しにくい状態がつくられている。そこは以前から「高電子密度層」(electron-dense layer)と名付けられてきた。だとすれば、この高電子密度層こそが何かの秘密を握っているにちがいない。そう、考えればよさそうなのである。
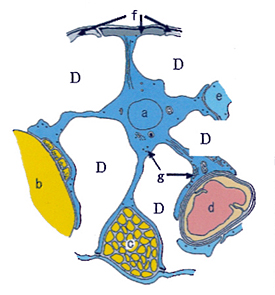
しだいに見えてきたことは、高電子密度層は大脳皮質の第1層と向かいあっているということだった。
なぜ、高電子密度層は大脳皮質の第1層と向かいあっているのだろうか。そこに錐体細胞の樹状突起があるからだ。錐体細胞は自分が皮質のどこにいるかにかかわらず、その基本たるべき樹状突起を第1層においていた。
樹状突起とはシナプスの受け側の構造である。シナプスは軸索の先端のシナプス終末で、ニューロンからニューロンへの信号の受け渡しをする。受け渡しはシナプス終末から放出されるニューロトランスミッター(神経伝達物質)によっておこなわれる。ニューロトランスミッターはシナプス間隙と水(細胞外液)とのあいだを泳いで、シナプスの受け側にある樹状突起のなかのレセプターに到達し、そのとき樹状突起に電気的刺激を発生させている。
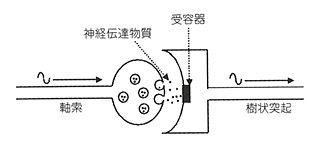
これまでは、おおむねそのように考えられてきた。ぼくもずっとそう思ってきた。しかしよくよく考えるとシナプスはけっこう面倒な操作をしているのである。ニューロンが運んできた電気信号を化学信号に変え、その化学信号が次のニューロンに到達して、そこで電気信号に戻るのだ。これはかなり面倒だ。しかし、すべての信号伝達をこのようにしなければならないという理由はないはずだ。
最終目的が樹状突起に信号が伝わるということなら、直接に電流を送ってもいいはずだ。ここにおいて中田仮説は新たな機能構造を提案する。グリアがつくる高電子密度層と錐体細胞の樹状突起とが構成する“小さな装置”がそこにあるのではないかという提案だ。もしそういうものがあれば、それこそがいままで見えなかった分配器にあたるものだろう。
中田はこれをELDER(Electron-dense Layer and Dendritic Ramification)と呼ぶことにした。グリアとニューロンの複合的信号伝達装置のようなものだ。ELDERとは、いわば水を神経伝達物質とする複合的シナプスなのである。
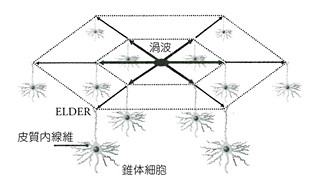
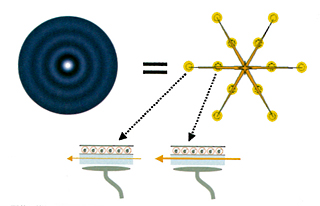
このアイディアは、かなり出来がいい。しかしながら、まだまださらに説明しなければならないことがある。ELDERがそういうものだとしても、そのスイッチは瞬時に入れたり切ったりできるようになっていなければならない。アセンブリーで水を入れたり出したりしているのでは遅すぎる。ここにおいて中田の仮説は、ニューロンを伝わってきた電気信号が消失することで発生する微量の熱がおこす渦波(vortex wave)に目をつける。ここからが、いよいよMRI研究者の面目躍如たるところになっていく。
脳には「渦流」があったのである。その渦流はどのように生じるかというと、均等に広がっていた水分子に乱流が発生して、局所的な凝縮状態がつくられ、その濃淡の差異によって渦波が生まれたのだった。
もう少し正確にいえば、渦波がELDERを通過すると、一時的にELDERの間隙の水が凝縮され、それがELDER間隙の誘導率を上げる。その瞬間にELDERに電流が流れ、次の瞬間に定常流に戻ったELDERは制止の状態になる。こういうことだろう。このようになっているのなら、ELDERはまさにさきほどから探していた分配器のはたらきをしていることになる。
おそらく、中心部から表面に流れる流体そのものは、もともとニューロンの活動に伴って生じる熱を効率よく逃す目的で発達したのであろう。無数のニューロンが情報処理のたびに発生させる熱量はそうとうなものである。そこで熱を逃す構造が必要になったのだ。いわば脳の空調機能というものだ。
ひるがえって考えてみると、中枢神経系の進化の手順はわかっている。「脊髄→小脳→大脳」の順にできあがっていった。小脳までの中枢神経はニューロンを中央付近において、その外側に電線を張るようにした。壊れやすいニューロンを保護するには適した方法だった。それゆえ中央付近で熱が発生し、ラジアル線維が残した「空気の通路」は空冷にはもってこいのものとなった。
一方、大脳はどうして自分自身を作ったのか。この空冷装置そのものを情報処理の機能にするように発達したにちがいない。そのため大脳ではニューロンが外側におかれ、電線が中心付近になるようにしたのであろう。このときELDERが活躍したはずだ。ELDERは皮質の表面全域に広がって、二次元の層をつくったのであろう。中田は、このようなELDERを含む層の全体をLGS(Lattice Gas Shell)と名付けた。脳はニューロンネットワークとLGSの二重構造をもったのである。

いろいろ省いてしまったが、だいたいは以上が2冊目の『ぷらす・あるふぁ』に述べられていた中田仮説の概要だ。ぼくはやっと斬新な仮説のおもしろさを感じるようになっていた。どきどきもした。
しかし、それでもいろいろわからないことが噴出もしていた。困ったなあと思ってはいたが、やむなくまたもや放っておき、別の脳科学の本をいろいろ漁っていた。くだらないおバカな脳関係の本が多くて困ったが、まあ、仕方がない。そして4年後、中田力の決定打ともいうべき本書『脳のなかの水分子』が登場したのだ。
今度も芦澤君のデザインであったが、まことにシンボリックに出来ていて、読みたくなるものがある。カバーにはMRIの写真を35カット型押しで並べてみせた。本文もタテ組になった。本文用紙がよすぎるのが気になるものの(マーキングがしにくいのだ)、あとは申し分ない。とくに文章がいい。ぼくはやっと気になってきた思考の便秘を脱出することになる。
本書はライナス・ポーリングのある論文との出会いの話から始まっている。ポーリングは化学賞(1954)と平和賞(1962)の二つのノーベル賞を受賞した生化学者で、量子化学者だった。オレゴン大学の出身だが、若き日にミュンヘンに行って量子力学を修めたので、ゾンマーフェルト、ニールス・ボーア、シュレディンガー(1043夜)が師にあたる。
ノーベル化学賞は、タンパク質の立体的二次構造をつきとめたことに対して与えられた。タンパク質は生命の基本であるが、それが生体機能をもつには、たんなるアミノ酸の連鎖から成熟して適度の立体構造をもつ必要がある。その最初の変化はペプチド結合による連鎖の特殊性に始まっている。これは二次構造で、アルファヘリックスという螺旋構造とベータシートという平面構造をもつ。ポーリングはそのあたりのことのほぼすべてを解明して、次のワトソンとクリックによるDNAの構造解明の準備を提供した。これがフツーの人名辞典にのっているポーリングの科学上の功績だ。
平和賞のほうは、大気圏核実験禁止協定を構想から締結にまでもちこんだことに対する栄誉であった。今日の核拡散防止条約(NPT)の前身にあたる。そういうポーリングだが、晩年はビタミンCの大量摂取療法を提案して顰蹙を買った。
しかし実は、脳の覚醒が脳における水分子の状態変化によって左右されていることを、最初に見抜いた天才科学者でもあったのである。
中田が注目したポーリングの論文は麻酔学に関するものだった。ポーリングは不活性ガスのキセノンに麻酔効果があることに関心をもち、しばらくそのことを研究していたのだった。
キセノンはどんな物質とも化学反応をおこさない。だから不活性ガスなのであるが、それなのにキセノンは麻酔効果をもっている。なぜなのか。そこでポーリングがいろいろ調べてみると、キセノンどころかどんな麻酔薬についても実は作用機序がわかっていないことを知った。まったく何たることか。
もともと全身麻酔はエーテル麻酔から始まっていた。やがてギリシア語起源の「アネスセーシア」という用語が麻酔にあてはめられた。直訳すると「不感覚」という意味をもつ。
すでによく知られているように、人間の「感覚」はニューロンを通して電気的信号に変換され大脳皮質に到達する。エーテル麻酔による「不感覚」は、この感覚処理の最後の部分、つまりは大脳皮質の機能を抑制することによっておこる。もっと正確にいえば、麻酔による「不感覚」はたんに感覚の麻痺だけがおこるのではなく、大脳皮質の情報処理機能のいっさいが抑制されることによっておこっている。これで「意識」さえ取り去られてしまうのだ。
なぜ、こんなことがおこるのか。一般に普及しているのはマイヤーとオーバートンが提唱した「脂肪溶解度説」というもので、麻酔剤は脂肪に溶けやすいものほど脳の中にいきわたるというふうに理解されてきた。しかし、これでは何の説明もしていないに等しい。それに対してポーリングはとっくに別の仮説をつくっていた。麻酔薬は水分子と水分子とをくっつきやすくしていると喝破したのである。全身麻酔効果のある薬剤が水のクラスター形成を安定化して、そこに小さな結晶水和物をつくりだしていると考えたのだ。

この仮説は当初は「水和性微細クリスタル説」と呼ばれていたが、いまでは「水性相理論」と呼ばれるようになっている。中田力はこの理論こそ、麻酔薬の効果の本質を解くだけではなく、人間の意識の根源が脳の中の水分子のふるまいに依存していることを告げているだろうことを確信した。
ニューロンには興奮性がある。活動電位のことをそう見立てるのだが、この興奮は正確にはイオンのはたらきによっていた。
ニューロンの内部を埋めている細胞内液はほとんどカリウムイオンで占められている。細胞膜を介して接している細胞外液はナトリウムイオンで占められている。二つのイオンのあいだには著しい濃度勾配ができあがっている。これはニューロンがつくりあげた勾配で、細胞内液からナトリウムイオンを汲み出し、そのかわりにカリウムイオンを取り込んでいる。これがイオンチャンネルというもので、ポンプ機能にあたっている。活動電位はこのイオンチャンネルがおこしている。
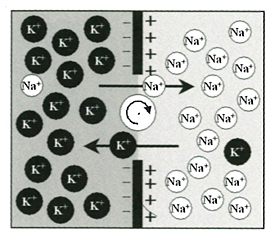

イオンは単独では存在しない。電解質が解けてイオンになれるのは、電解質の分子をつくっている二つの原子や原子群のあいだに水分子が入りこむからだ。水分子がいなくなると、電解質はもともとの分子に戻ってしまう。
その水分子も自分自身のつくる水に溶けてイオン化する。その結果、水素イオンと水酸化物イオンができる。この水素イオンにも特異なチャンネルがある。プロトンチャンネルというもので、ミトコンドリアの内膜で生体エネルギーを取り出すのに重大な役割をはたしている。
以上のナトリウムチャンネルもカリウムチャンネルもプロトンチャンネルも、すべてイオンが通る道である。このようなチャンネルが互いに結びあっている構造空間では、外にも内にも「水」がある。これが細胞外液と細胞内液になる。しかし、これだけで生命のチャンネルが仕上がっているなら、麻酔は効かないだろう。ということは、このチャンネルが積み重なっただけで「意識」が生じたわけではないはずなのだ。
では、ほかにはチャンネルは考えられないだろうか。そういうチャンネルがあるとしたら、イオンではないがイオンのように小さな分子でなければならないのだが、そんなものがありうるだろうか。
ありうるのである。それは「水分子」そのものなのだ。このことは長らく仮説のままにあった考え方だったのだが、最近になって水分子だけを通過させるチャンネルが発見された。水分子に特異なチャンネルが見つかったのだ。それがアクアポリンだったのである。この発見はピーター・アグレたちによるもので、2003年のノーベル賞の対象になった。中田の渦理論は一挙に構想を膨らませていった。
高等動物にとって大切なことは、生体内の環境を一定に保っておくことにある。ホメオスタシスの維持である(211夜)。鳥類と哺乳類はなかでも体温を一定にするため苦労した。
ぼくは平温が36度5分以上ほどあって、しばしば握手をすると「おや、手が温かいですね」と言われる。それが38度以上になると、全身がだるくなる。38度5分になってもへいちゃらな連中がいることは承知しているが、ぼくはからっきしダメである。てきめんに衰弱する。
ともかくも人間というもの、そのくらい温度に敏感な生体システムなのである。なぜこんな正確な温度調節が必要なのかというと、細胞のなかでおこなわれている代謝に関係がある。ここで酵素による代謝の促進が進んでいる。それを維持するために厳密な温度が調整される。そういう熱化学が必要なのだ。
ふつう、多くの化学実験は室温程度があれば十分に進行する。25度程度で十分だ。ところが人間の体温が25度になっては生きてはいけない。なぜ、われわれはこんな厳密な温度管理のなかにいることにしたのだろうか。中田仮説の回答は明快だ。それは「脳を管理するため」だったからではないか。脳神経系の進化にとって厳密な熱化学的な温度管理が必要だったのではないか。
この管理に大きな役割をはたしたのが「脳のなかの水分子」たちだったのだ。水は熱の伝導にすぐれていて、かつ保温力が高い。それだけでなく流体であるのだから、どんな形態にもなじむ。中田はこのように書いてきて、次のように結んだ。「ここから、脳の渦理論とよばれるものが生まれてきた。その主役は水だったのである。脳は、水を管理することでニューロン・ネットワークを保護する発泡スチロールのような緩衝材を作り上げ、同時に、熱の流れを大脳の機能素子として利用することに成功したのである」。
以上が、本書をまことに乱暴に単純化してしまった要約だ。1冊目、2冊目との関連が多かったので、こんなふうな要約にした。
ぼくが理解していった手順だけを書いたので、わかりにくかったかもしれないが、詳しいことを知りたいなら、本書を手にとり、すぐにカフェに入ることである。では、最後になったけれど、あまり知られていないかもしれないことを書いておく。MRI(磁気共鳴画像)とは何かということだ。
MRIは何をあらわしているかというと、びっくりしないでほしいのだが、実はまさに水分子の画像をあらわしているのだ。体のなかを作っている水分子が、与えられた特別の周波数に音叉のように共鳴しておこす信号をとらえて画像にしたものなのだ。それがMRIなのだ。
もともとMRIの基本となった物理現象はNMR(核磁気共鳴)というもので、ブロックとパーセルが発見して1952年にノーベル賞をとっている。原子核が磁場におかれたときに示す共鳴現象のことである。原子核はスピンという回転運動をしているのだが、重力などの影響でわずかな歳差運動をおこす。構造が完全対称性をもっていない原子核は、このときよろめいてしまう。これをラーマー歳差運動という。このラーマー歳差運動をする原子核を磁場の中におくと、歳差運動の周波数に従った共鳴現象を呈するわけだ。
その後もNMRはコンピュータ技術の革新にともなって、フーリエ変換法が導入されたり、それが画像医学という新分野に発展していったりした。そしてNMRを発展させたMRIは、水分子の水素原子核が示す共鳴現象をとらえることになったのである。
MRIは水素原子核からの信号を受け取っている。水分子の水素原子核は磁場の中では上を向くか、下を向くかのバイナリーな行動をとる。水分子は互いがぶつかりあってエネルギーの交換をしつづけている動態系なのだ。
あらためて言うまでもないだろうけれど、われわれもまたそのような水分子をもった動態系である。その動態系が磁気共鳴を得るには、われわれは大きな磁石の中に入る必要がある。MRIで患者が大きな円筒磁石の中に体を入れるのはそのためだった。「はい、息を大きく吸って、はい、そこで止めて」。
MRIとは水分子の奏でる生体のシンフォニーなのである。中田力が「脳のなかの水分子」に注目できたのは当然だったのだ。そのうち、新潟大学の統合脳機能センターを訪れたい。そのときは、中田さん、いろいろのことを教えてくださいね。